



| 孟小龍:一個癌症機制的新假說 |
| 送交者: mxl 2011年12月18日20:22:10 於 [健康生活] 發送悄悄話 |
|
一個癌症機制的新假說 孟小龍
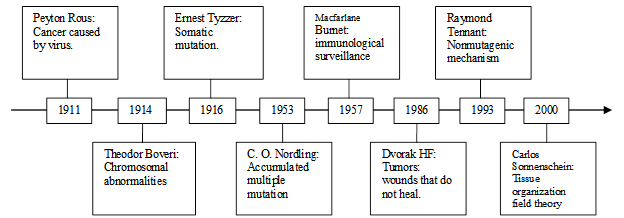
摘要:對一些資料的研究使我們導出了一個癌症機制的新假說。首先,癌症僅現於那些有複雜的損傷修復能力的多細胞生物。其次,損傷作為危險因子可見於臨床所有癌症。 最後,致癌基因的活化不僅見於癌症,也見於正常的生理和非癌症病理過程。我們所提出的假說認為癌症是一個自然的損傷修復相關的過程,包括致癌基因的活化,細胞因子的分泌,幹細胞的調集-分化和組織重塑。損傷激活了某些細胞的致癌基因,而後者分泌細胞因子來徵募幹細胞以癒合損傷。然而,如果損傷的病因或損傷持續,例如在持續的UV和致癌物暴露下,連續的損傷修復過程會導致臨床癌腫。當損傷存在時,自然界沒有一個系統來停止或反轉處於中間狀態的損傷修復過程。癌症機制的結果或者是癒合損傷,或者是系統耗竭(死亡)。這個癌症機制的邏輯與機體其它生理代謝機制一致—為了生存。這個假說幫助理解許多突變理論所致的癌症謎團,例如為什麼癌症僅現於一小部分多細胞生物,儘管它們在DNA複製時都有潛在的突變危險。這個假說可以用於解釋和指導癌症預防,復發,轉移,體外體內研究和個體化治療。 此文的英文原版發表在 Cancer and Metastasis Review 期刊的2011年12月16日網絡版,在此譯成中文與中文讀者探討。文中的圖表僅部分譯成了中文。文獻請參閱英文原版:http://www.springerlink.com/content/lx671t71467n172k/fulltext.pdf 1 背景:當前的癌症理論及疑問 由於癌症尚未能治癒,自從上世紀初以來,許多假設或理論一直不斷被提出(圖1),包括癌症由病毒引起[1],染色體異常[2, 3],體細胞突變[4],積累性多步突變[5],免疫監視[6, 7],不愈損傷[8],非突變機制[9],和組織協調域理論[10]。
圖 1. 癌症假說或理論的時間表。
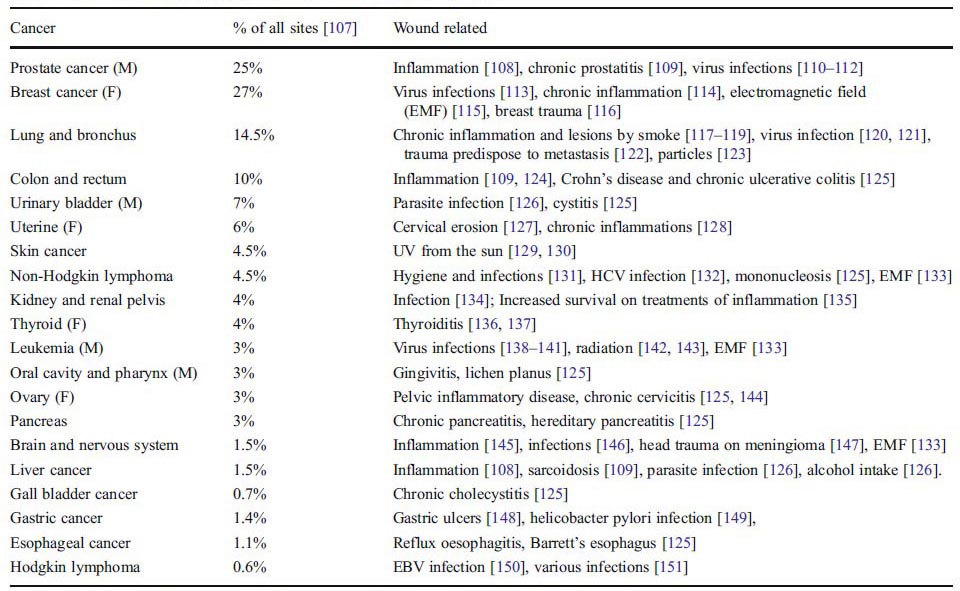
目前占優勢的癌症理論認為癌症是非控制的體細胞繁殖,是由控制細胞生長和分化的一些關鍵基因的進展性的累積突變引起的[11–13]。免疫監視理論是突變理論的一個補充。當突變成功逃逸免疫監視後,臨床癌症就會形成[7]。然而各種有關突變理論的自相矛盾的情況時有報道,比如癌細胞的繁殖速度低於正常細胞,癌的悄悄轉移,睾丸癌細胞注入正常芽胚後生成正常組織,正常鼠卵組織注入脾後生成癌組織,癌自然轉歸等[10, 14]。此外,突變理論還不能解釋下列現象: 1.如果一個突變就引起癌症,得癌的機會就太大了。突變需要細胞的分裂。成人體內每天有1012個幹細胞分裂[12]。如果每個細胞分裂的突變率是1.1×10-8 [15],且癌症由一個突變引起[3, 16],那麼理論上每個人的癌症發生率會極高:每天1.1×104點突變或癌症(1012幹細胞分裂×1.1×10-8點突變/細胞分裂),意味着每人每天都會得癌。這顯然不符合實情。於是,多步突變理論(突變-潛伏-突變…致癌)[5]和DNA 修復理論便產生了,以符合實際的癌發生情況[12]。 從另一個角度看,如果癌症是由多個特定突變所致,則得癌的機會又太少了。如果一個癌起因於5個特定點突變[11],理論上的癌發生會是每人每天1012×(1.1×10-8)5=1.61×10-28,或者對一個120歲的人來說是1012 ×(1.1×10-8)5×365×120= 7.05 ×10-24。這相當於在1.42×1023個人中有一個人會得癌, 也就是說全世界沒人會得癌。臨床上報道散發性的結腸癌有5-10個特定的基因改變,甚至每個細胞可以達11,000個基因改變[17–19]。這便產生了一個疑問:突變究竟是癌症的起因還是結果?類似地,如果一組病人有相同的基因突變或表達同樣的蛋白(比如骨腫瘤細胞系CDKN2A的缺失[20];小細胞肺癌的p53和Rb突變[21])或者不同癌症有同樣的突變(比如各種癌症的同源性磷酸酶-張力蛋白改變[22]),理論上隨機突變導致這些癌症同時發生的可能性幾乎不存在。 2.突變理論難以解釋在不同機體中引起癌症的時間差異。在兩年之內,多於50%的Sprague–Dawley大鼠會自然發生癌症[23],而人在兩歲之內這一發生率遠小於百萬分之一[24]。人和大鼠骨髓幹細胞的倍增時間分別是25.2[25]和31.5小時[26]。人細胞的分裂時間並不比大鼠細胞慢,這意味着人細胞的DNA複製錯誤的機會並不比大鼠的少。單從突變理論來看,人沒有理由在相同的DNA複製時間內顯示比大鼠低得多的癌發生率。相似地,累積突變也不能解釋為什麼癌症的發生並不與不同物種的壽命相關,比如千年的無癌古樹對比12天的有癌果蠅,儘管它們有相同的DNA複製機制[27, 28]和相似的致突變的環境危險。此外,癌症僅存在於某些多細胞生物而免於另一些多細胞生物的事實,表明突變不應該是癌症的必要前提,因為所有的多細胞生物在DNA複製中都有突變的風險,而僅有其中一小部分有癌症。 3.兩個種屬的多細胞生物細胞在相同潛在突變環境中有不同的癌化結果。有許多血吸蟲相關的人膀胱癌和前列腺癌的報道[29],而並沒有發現血吸蟲自身得癌,儘管它在相同的癌環境中有相同的潛在突變危險。如果血吸蟲相關的膀胱癌是由人p53突變引起[30],為什麼相同的環境從不襲擾血吸蟲的p53基因[31]並在血吸蟲身上發生癌症呢? 4.癌症的復發也不能被突變理論解釋。理論上,如果有一個活癌細胞在外科,化療,放療後存活,另一個有1x1012細胞(約1公斤)的癌腫會在80天后形成,如果細胞的倍增時間是48小時的話[32]。如果所有的癌細胞都被標準殺滅了,且新的癌細胞又是由累積突變產生的話,根據突變理論[11],象第一個癌產生所需的時間一樣,又得幾十年,而這與臨床病例不符,如多數乳腺癌在5年之內復發[33]。 5.無論是突變理論還是免疫監視理論都不能解釋癌症發病率在鼠(>800 天) [34]和人(>85 歲) [24, 35, 36]極老年之後翻轉。如果突變和免疫監視或DNA修復的失敗是癌症的原因,那麼在極老年體內的衰老細胞會有更多的機會得癌。對此發病率翻轉的一個解釋是自然選擇使得癌症易患人群難以生存—老年存活者不易得癌[36]。然而這一機制,如果存在的話,卻與為什麼此現象不存在於其他的老年性疾病(比如心臟病)[37]相矛盾。 2 新癌症機制的推導 對於基因突變與癌症的關係,有兩種可能性:1. 象突變理論所述,基因突變是癌症的起因。2. 取而代之,基因突變是癌化的中間過程或結果。由於有如此多的與突變理論不符的現象,應考慮更好的癌病因學說。 2.1 損傷作為癌症的危險因素 癌症伴隨着致癌基因活化,而後者也參與損傷和損傷修復過程。如果損傷被定義為由物理的(放射線,電磁場,外傷,顆粒等),化學的(致癌物,毒性化學物,重金屬等)以及生物的(炎症,微生物感染,自由基,營養缺乏,老化,緊張等)破壞而引起的細胞死亡,損傷作為危險因素幾乎可見於臨床所有的癌症,諸如慢性炎症和前列腺炎對於前列腺癌;病毒感染和外傷對於乳腺癌;吸煙引起的組織損傷對於肺癌;慢性潰瘍性結腸炎對於結腸癌;UV損害對於皮膚癌;以及病毒感染,放射,電磁場對於白血病(表-1)。實驗顯示儘管Rous肉瘤病毒在血液中循環,但它只在損傷炎症處引起腫瘤,且抗炎藥可抑制腫瘤生長[38]。另一項研究甚至顯示帶有炎症基因,LPA和ATX的轉基因小鼠有較高的乳腺炎以及隨後而致的乳腺癌發病率,此與臨床情況相符[39, 40]。這些損傷與癌症的廣泛相關提示了兩種可能性:損傷引起癌症,或者癌症應付損傷。
表1. 人的癌症和作為癌症危險因素的損傷
百分數源於文獻[107]中估計的2009年的新病例。M 代表男性,F代表女性,無特別指出代表兩性。最後6行由該文獻表1算出,其他百分數源於該文獻圖1。
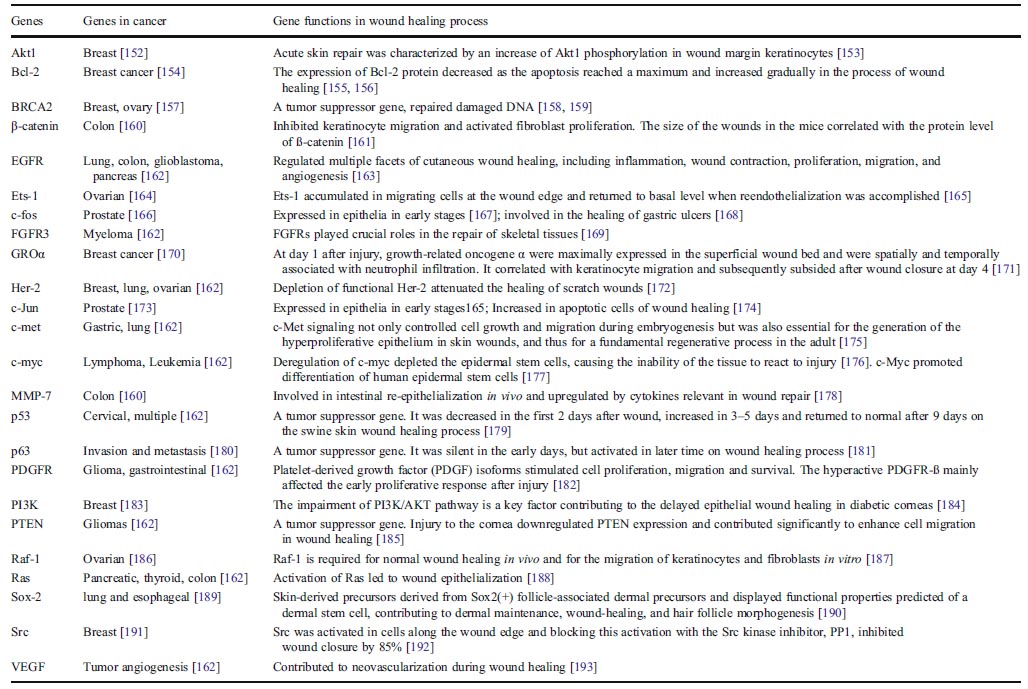
2.2 損傷修復中的致癌基因 如果致癌基因被定義為存在於正常細胞[41],且過度表達時(抑癌基因相反)可使正常細胞轉化為癌症細胞的基因[42–44],在分子水平,許多(如果不是所有)在癌中發現的致癌基因也都活躍在早期的損傷修復過程中以增殖修復細胞。在癌中失活的腫瘤抑制基因在損傷修復早期失活,而在修復晚期復活以停止修復細胞的繁殖(表-2)。這些致癌基因在損傷修復中的活動提示致癌基因機制在損傷修復中也扮演重要角色。如果癌症是致癌基因過度表達的結果,很可能癌細胞作為活化致癌基因的組裝體在損傷修復中起作用。一項荷瘤小鼠的血漿癒合傷口比正常小鼠血漿快的研究支持這一假說[45]。 表2。致癌基因和抑癌基因見於癌症和損傷修復過程
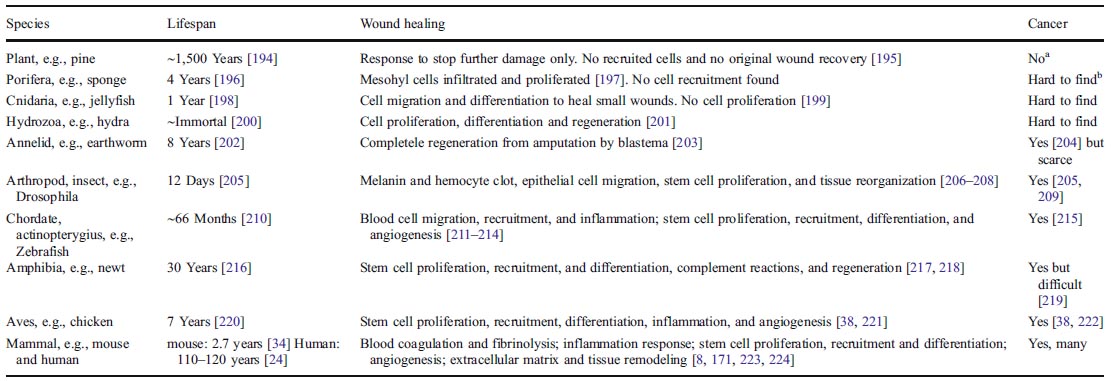
2.3 癌症在多細胞生物中的出現 拓展至更廣闊的視野,多數地球上的生物沒有癌症,儘管它們都有相同的NDA複製機制(表-3)。從簡單到複雜的物種,損傷修復過程也從最簡單(無原傷口複合)到中等複雜(細胞遷移,分化和再生),到最複雜的損傷修復過程(包括前者再加上炎症反應,幹細胞徵調,和組織重塑等)。癌症的發生與損傷修復的複雜程度相符。越複雜的生物,功能和特殊分化的組織就越多,損傷修復就越複雜。在某種損傷修復的複雜水平上,尤其是帶有炎症和調用幹細胞(非再生)過程的生物,癌症便出現了,無論其壽命長短。這個現象提示自然對複雜的物種已選擇了一個共同的損傷修復機制,這個機制與癌症有關。 表3. 有與沒有損傷修復能力的多細胞生物和癌症
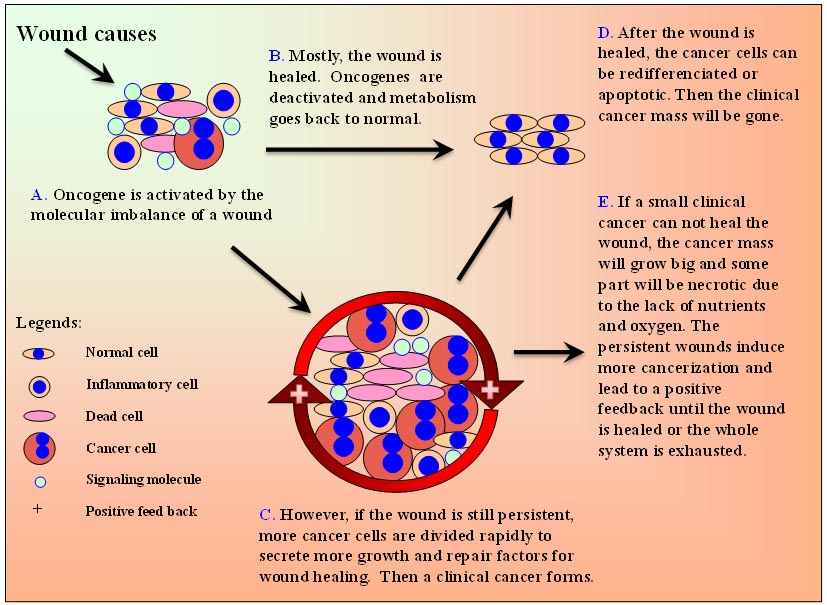
a 植物蟲癭(也叫“植物腫瘤”)是植物(宿主)與土壤桿菌屬[223],真菌[224]和昆蟲[225]的寄生交互作用。土壤桿菌插入一段DNA至植物細胞,這與源於哺乳動物正常細胞的癌細胞不同。 b “難於找到”的意思是這一條在網上難以找到,標誌着這一情況少見,且未被注意或集中的研究過。 以上所有損傷與癌症之間在進化,疾病以及分子水平上的關連強烈表明一種可能性:損傷信號分子使已有的致癌基因(和其它一些基因)的過度表達,導致某些染色體和癌細胞表型變化[42],而所有正常細胞的致癌基因(不限於致癌基因)對損傷的反應僅是細胞為了生存的自然代謝的一部分。因此,可以合邏輯地導出一個新的癌症理論:癌症的形成是一個生物用它來修復損傷的自然手段。 2.4 哺乳動物的癌症機制 下列損傷-致癌基因-損傷修復(WOWH)的情景被用來描述哺乳動物的癌症機制(表-4, L1-10)。當上述定義的損傷發生在哺乳動物時,機體便開始了複雜的,有炎症和幹細胞參與的損傷修復過程(L1)。生長因子,細胞因子和其它從損傷區域細胞來的蛋白分子破壞了正常的分子代謝(L2),導致相應的致癌基因活化(表-2)及誘導某些細胞(幹細胞或分裂活躍的細胞)癌化(L3)。致癌基因活化的細胞能分泌分子來調用幹細胞,刺激幹細胞繁殖以及增進細胞分化以修復損傷(L4,圖2A)。致癌基因在損傷早期被活化,而抑癌基因在晚期或損傷癒合後被活化(L5)。多數情況下,經上述努力之後,損傷被癒合了。致癌基因失活,抑癌基因復活,代謝又回復正常(圖2B)。然而,如果損傷仍持續(例如在持續的UV和致癌物的暴露,以及慢性損傷的情況下),這個WOWH機制會繼續,致癌基因會持續活化,更多的癌細胞(過度活化的致癌基因使正常細胞轉化為惡性細胞[43, 44])會分裂以分泌更多的修復損傷的分子,導致臨床癌症的形成(L6,圖2C)。損傷癒合後,癒合環境的分子會啟動癌細胞的分化(L7)或凋亡(L8)。結果臨床癌塊會消失(圖2D)。然而,如果一個小的癌症不能修復損傷,癌塊會長大,且由於缺乏營養和氧氣,癌症自身的某些部分會出現壞死(包括炎症---新的損傷)(L9)。原來的和新的損傷,兩者都會引起更多的癌化過程,進而引起一個正反饋,直至損傷癒合,或者整個系統耗竭(L10,圖2E)。 表4. 癌症機制假說的邏輯
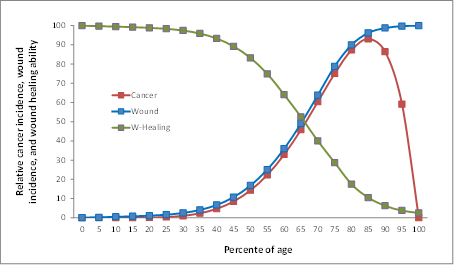
圖 2. 損傷-致癌基因-損傷修復機制示意圖 3 WOWH機制的進一步解釋 3.1癌症機制對於生存的兩難 所有上述WOWH過程都是由損傷細胞,炎症細胞,癌細胞,幹細胞,細胞外基質細胞以及癒合的細胞間的分子反饋來精細調節的。從損傷和炎症細胞分泌的信號分子越多,癌化細胞的響應就越多。從癒合的損傷分泌的信號分子越多,則越多的癌化細胞停止工作(表-4,L1-10)。不同的損傷激活不同的致癌基因,誘導不同的癌以產生不同的分子來修復損傷。這一WOWH機製發展的非常之好,但除了一個缺陷:如果損傷持續(例如慢性微生物感染,慢性子宮頸糜爛等),癌塊會越長越大,而我們的系統中沒有一個自然機制來阻止那個正反饋環直到整個系統耗竭(死亡)。正常細胞在發育,損傷修復以及這些過程後自我平衡的邏輯是可以理解的[46]。然而,如果損傷持續,自然應該選擇什麼邏輯來修復損傷呢?面對持續的損傷,自然選擇除了與損傷抗衡至系統耗竭之外,在邏輯上似乎沒有其它選擇,因為晚期癌症自然恢復的機會確實存在。更重要的是,在某一點,比如在癌症晚期,打破這一正反饋,意味着所有對損傷的分子反應都需要停止或反轉,導致生存的邏輯問題—對粘膜,皮膚,肝臟,血細胞和其它快速代謝的組織的損傷和細胞老化無反應或逆向反應。一項研究顯示試圖產生過度表達野生型p53基因的小鼠總是在胚胎發育上失敗,而部分表達p53的小鼠雖表現出抗自發腫瘤的功能,但它們的生命卻顯現出早期衰老,包括壽命減少,骨質疏鬆症,普遍的器官和組織萎縮,遲緩的損傷修復,以及對緊張的耐受降低[47]。 3.2 在非常老年的癌症發生率翻轉 當癌症被認為是對損傷的功能性修復組織後,癌症發生率在極老年時翻轉之謎便可以解釋了:創傷,炎症,感染,毒素和細胞老化的積累以及損傷發生率隨年齡的增加而增加[48]。由於細胞的老化,損傷修復能力隨年齡而下降[49, 50]。癌症,作為功能性的損傷修復組織,其發生率由於損傷發生率而隨年齡增加,卻由於在極老年時損傷修復能力的衰竭而減少(圖-3)。女人比男人的癌發生率低[24],也可用女人的損傷修復能力較高來解釋[51]。
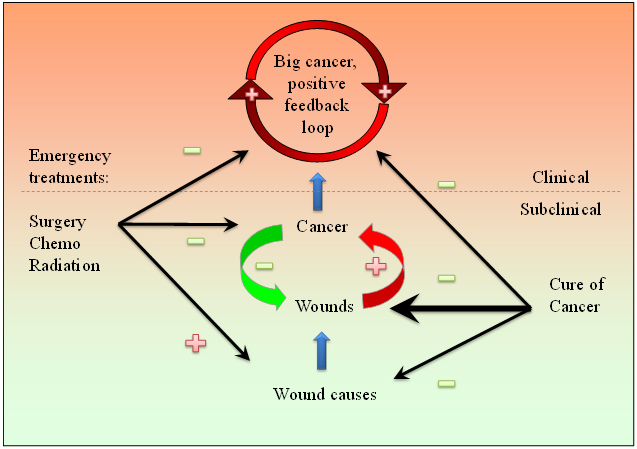
圖-3. 哺乳動物的損傷發生率(藍),損傷修復能力(綠)和癌症發生率(紅)與年齡的關係。損傷發生率隨年齡上升而損傷修復力隨年齡下降。癌症作為功能性的損傷修復組織,其發生率由於損傷的增加而隨年齡上升,但由於在非常老年時衰竭的損傷修復能力而下降。所有的數據點都是根據參考資料24,190,191和192而描述的。 其它上述不可被突變理論解釋的現象可以被WOWH機制解釋:(1) 臨床患癌症的機率不是取決於基因突變的數量,而是取決於潛在的損傷和損傷修復力。多個致損傷因子會誘導多條癌化通路來對抗損傷。(2) 種屬間癌症發生的時間差異是由於種屬間的細胞老化的時間不同。老化的細胞環境是哺乳動物的生物損傷的原因之一。WOWH機制會隨着哺乳動物壽命中越來越多的衰老細胞而啟動,而不是突變的積累時間。(3) 不同的生物在相同的潛在致癌環境中有不同的癌化結果是因為它們有不同的損傷修復機制。沒有象哺乳動物那樣的WOWH機制的生物,在突變環境中就不得癌。(4) 癌症復發的快慢取決於潛在的損傷和個人損傷修復的能力。持續的損傷(可能來源於緊張[51],飲食[52],空氣污染[53]等)會加速復發的腳步。 3.4 修復機制而不是修復錯誤 損傷修復和癌症的共同點提示了兩個可能性:一個是這一過程錯誤地處理損傷,導致不癒合的損傷, 或者癌症[8, 54]。另一個是大自然發展了一個過程來修復損傷——總是伴隨損傷的WOWH機制(圖-2)。第一個觀點與上述的正反饋情形相符。然而從更廣一些的視野來觀察癌基因的活動,如在妊娠[55, 56],胚胎發育[57, 58],月經[59],骨和牙的發育[60, 61],以及損傷癒合中(表-2),那些致癌基因活動的共性提示帶有相同功能的分子在正常和癌症狀態中都有,而不是在癌症中重新開始的錯誤程序。基因突變不僅存在於癌細胞中(以較高的發生率),也存在於良性增生或癌前病變(以較低的發生率)[19, 62, 63],提示癌症的基因突變應該是組織適應的結果,而不是癌症的起因。癌症僅現於損傷癒合相對複雜的物種,以及女性的癌症發生率約是男性的一半的現象[24],也提示癌症不是從一個簡單的突變或錯誤程序而來。在所有這些方面,WOWH機制的解釋更恰當。只有把癌症看作一個主動的損傷修復組織,上述的癌症之謎和下面的應用才能更好的被理解和統一在一起。荷瘤小鼠的血漿癒合損傷快於正常小鼠血漿的研究[45]是癌症分泌具有癒合體內損傷能力的分子的直接證據。 4 WOWH機制的應用 4.1 癌症治療 區分癌症作為功能性還是錯誤的組織的意義在癌症治療的戰略上十分重要。如果癌症是一個細胞程序的錯誤,殺死癌細胞是治療此病的唯一選擇。然而如果癌症是損傷修復過程的一部分,僅僅殺癌細胞可能不會導致痊癒。前面一個癌塊拿走了,新的一個又會長出來[64]。前面一個信號通路被抑制了,新的一個又會被激活[65]。只要潛在的損傷存在,WOWH機制就會對它有反應,直到損傷修復力衰竭(死亡或老到不反應)。此外,信號通路中的分子不是彼此單線聯繫的。它們每一個都在信號網絡中同時聯絡許多其它分子[66]。阻抑一兩個網絡的節點可以暫時延遲信號流,但不會徹底阻斷它,只要損傷還存在。換句話說,摧毀癌細胞而沒有修復潛在的損傷會最終導致復發,因為大自然發展的癌症機制在活的哺乳動物身上不能被摧毀。這也可能是為什麼最常見癌症(肺癌,乳腺癌,前列腺癌和結腸癌)帶有轉移的病人的生存率在過去的幾十年裡都沒有顯著改變的原因[67]。因此,治療癌症的戰略應該是在目前的標準治療(手術,放療,化療)打破那個正反饋環後,專注在潛在損傷和分子平衡的治療上(圖-4)。未知的潛在損傷和個體化的分子不平衡使得以此治癒癌症比傳統的標準治療要困難得多,但那卻是治癒癌症的唯一通路。它不但有賴於主要藥物治療,還會受制於其它可能影響損傷癒合的因素,比如緊張,飲食,環境,生活方式等。癌症的治癒是一個在分子水平上的個體化的多維自我平衡過程(圖-5)。依據對WOWH機制的理解,可以解釋癌症的自發性消退,且它可被認為是正確的整體分子平衡而治癒癌症的極端例子。晚期創傷液較早期創傷液減低移植腫瘤在小鼠體內的形成,這提示來自癒合的損傷的分子舒緩了癌症的負擔[68]。據此推測,很可能來源於自體癌腫的分子能預防或治療原發癌的復發。以下的研究支持這一假說:從腫瘤細胞來的侶伴蛋白抑制幾種小鼠模型的腫瘤生長[69];第二個移植瘤抑制小鼠第一個移植瘤的生長[70];以及EGF-癌症疫苗減少炎症[71]。
圖-4. 圖示損傷,癌症,反饋與治療。物理的,化學的和生物的因素引起損傷。癌症對損傷反應而癒合它。如果損傷的刺激持續,正反饋會形成損傷癒合或全身耗竭。 手術,化療和放療能夠快速停止正反饋環,但這些治療本身也是損傷的誘因。 如故損傷持續,另一個癌症會復發或誘發。為了治癒癌症,所有三個方面,正反饋環(癌腫塊),損傷和損傷誘因都得顧及,儘管後兩者更加複雜和困難。+ 表示增長,促進或正反饋。- 表示下降,減少或負反饋。
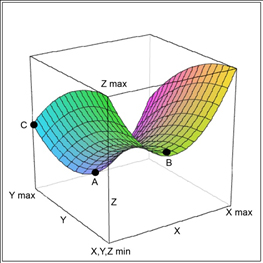
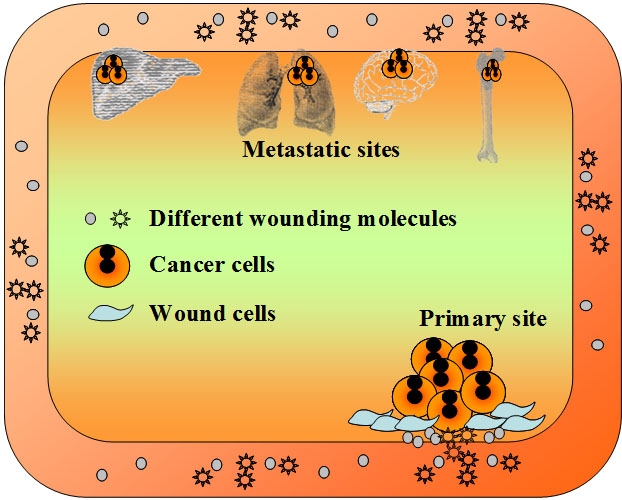
圖-5. 疾病,包括癌症治療的分子調節。X軸:主要治療分子的計量。最佳計量為最大和最小之間(B點)。 Y軸:所有其它可能的分子調節在癌症治療中的影響,從藥物,食物,生活方式到心理影響。 最大Y表示最佳影響,最小Y表示最壞影響。Z軸:疾病,癌塊或癌化過程。最小Z表示無疾病,而最大Z表示疾病的最壞狀態。A點:無疾病(癌症)點,由最佳X和最大Y達到的。 僅在這種情況下疾病(癌症)可以治癒。B點:部分臨床響應點,出於最佳X但是最小Y(正確的主要治療,但不幸的是有錯誤的生活方式等的影響)。C點:出於最大Y但是最小X的部分臨床響應點(錯誤的主要治療,但有正確的生活方式等影響)。Z=X²-Y²意味着治療結果有賴於多維的分子平衡過程。癌症的自發性消退可被認為是在這個模型中達到A點的極端例子。 4.2 癌轉移的解釋 癌轉移被認為是癌細胞從原發部位到遠處組織的遷移[72]。但是不清楚為什麼許多細胞系在動物模型上不發生轉移[72–74]。此外,癌細胞如何從肺,乳腺和皮膚跨越血腦屏障轉移至腦不清楚,因為似乎只有癌細胞能跨越血腦屏障, 而小得多的淋巴細胞卻不能[75]。基於WOWH機制,另一種“無細胞遷移的轉移”是可能的:相應的遠處癌基因可以被持續循環的損傷分子所激活,這些損傷分子來源於損傷以及癌症相關的壞死/炎症。有持續癌基因激活的細胞會轉化和過度增殖,導致遠處另一個癌腫的形成——可被認為是“廣義轉移”(圖-6)。例如,PI3K/AKT信號通路在乳腺癌[76]和骨再造中[77]都被激活。循環中激活乳腺癌細胞的PI3K/AKT基因的分子,也會刺激正在活動的骨細胞的相同基因,導致PI3K/AKT的過度表達和骨細胞的異常增殖——無細胞遷移的廣義骨轉移。一項第二個腫瘤誘導的研究[38]是很好的證據支持這個廣義轉移的假說:在小雞上引起損傷腫瘤的Rous肉瘤病毒不會引起腫瘤轉移。然而,遠離第一腫瘤的損傷會在第二損傷處發展另一個腫瘤。甚至某些損傷相關分子(TGF-β, αFGF和bFGF)可以替代實際的損傷而發生第二個腫瘤,提示啟動第一個損傷腫瘤的相似分子在第二損傷處激活了損傷相關細胞的癌基因而發展成了第二個腫瘤——無腫瘤細胞遷移的廣義轉移。從廣義轉移的概念的角度,容易理解為什麼在從原發灶到遠處轉移的途中難以找到連續的癌細胞落腳點,以及轉移灶如何跨越血腦屏障出現在腦中。只要損傷分子在體內循環,根據WOWH機制,它們會促進那些在生理代謝中有相應癌基因活動的組織中的癌基因活化,尤其是如果在那裡有損傷修復過程。
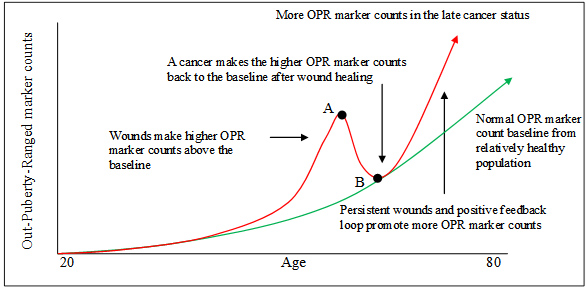
圖-6. 廣義轉移。遠處組織中有生理代謝活動的相應的致癌基因可被持續在循環中來自損傷和癌症相關炎症的損傷分子所激活。有持續激活的致癌基因的細胞會轉化和過度增生,導致遠處組織在沒有原位癌細胞遷移的情況下的另一個癌腫。 4.3 腫瘤標記物的波動及應用 在癌症中由於癌基因和其它癌相關基因的活動,許多蛋白質分泌到癌細胞外並進入血循環。由於血清樣本比組織樣本容易獲得,研究人員一直追蹤着癌症狀態的血清標記物[78, 79]。然而到目前為止幾乎沒有標記物被證明臨床有效。在一個研究中可預測的標記物,在其它的研究中則可能無效[80, 81]。在癌症中,生物標記物的臨床應用困難是由於理解它們的問題。如果認為一個標記物是癌症癌組織的唯一標籤,就找不到這樣的標記物用於癌症篩選,因為在癌症早期的所有標記物都可見於其它非癌的損傷狀態[82] (表-2)。基於WOWH機制,損傷,癌症和損傷修復是相互作用的。損傷促進癌症,而癌症修復損傷(圖-2,4)。損傷,癌症和損傷修復共享大多數共同分子。僅僅在癌症進入正反饋環時,基於癌症的損傷才會激活一些少見於非癌狀態的分子通道。因此,很難找到癌中高特異性的標記物。另一方面,損傷,癌症和損傷修復是一個動態過程。損傷刺激癌症相關標記物的表達(表-4,L2,L3),而癌症可能減少那些表達如果損傷狀態有所改進的話(表-4,L7,L8)。新的損傷,癌症治療,飲食和生活方式[51–53]會影響損傷-癌症狀態(圖-5)進而改變癌症相關標記物的表達,即使在癌症的晚期。所以,不可能用一種癌症狀態的標記物來預期後來癌症狀態的結果。這就是為什麼原發癌症的組織標記物與後來的轉移期有些不符的原因[83]。再者,不同的損傷激活不同的癌基因,誘導不同的癌以產生不同的分子癒合損傷,儘管這些損傷和癌可能發生在同一器官。因此,一個標記物改變不能覆蓋所有由組織或器官命名的癌症的損傷-癌症情況,如乳腺癌的ER±, PR± 和 Her-2±[83]。 基於WOWH機制,可以推測另一個癌症相關標記物的變化並用於臨床(圖-7):如果在20歲時的癌相關標記物陣列的每一個平均值±2標準差都設為青春值(成人的參考範圍),年紀大的人會有越來越多的超青春值(OPR, 在20歲時的平均值±2標準差之外)的標記物隨年齡增長,因為累積損傷和老化細胞隨年齡而增多[48]。然而由於癌基因活化,OPR在相對健康人群的計數(圖-7,綠線)要比癌前病變以及癌轉移的人(圖-7,紅線)低。損傷-損傷修復的相互作用,癌基因活化以及癌細胞的分泌會產生一個OPR標記物計數的峰。損傷癒合後,增高的OPR標記物計數會回落到正常基線上。這就是為什麼在癌症早期,標記物可能高或不高,比如結腸癌和乳腺癌的CEA和CA15-3[84]。如果損傷是間歇性的(如在每年假期中UV暴露的情況下),OPR標記物會上下波動。這可能會造成許多臨床“假陽性和假陰性”的判斷或研究中矛盾的結果[85, 86],而這些是由於對人群的取樣時間所造成的(圖-7,A,B點)。如果損傷持續或癌症進入了正反饋環(如癌症晚期),更多的損傷分子和對應基因會參與進來[84, 87, 88],OPR標記物計數會維持在高水平。通過監測動態OPR標記物計數與正常基線的比較,可以看出體內的癌化狀態,且可以指導個體化治療朝着正常基線的方向進展至癌症痊癒。
圖-7.癌症相關生物標記物與年齡,損傷和癌症的相互作用。20歲時的癌相關標記物陣列的每一個平均值±2標準差都設為青春值(成人的參考範圍)。超出這個平均值±2標準差被設為青春值外(OPR)。由於損傷積累和細胞老化,相對健康人群的正常OPR標記物計數隨年齡上升(綠線)。損傷使OPR標記物計數高出基線,而癌症在損傷癒合後使較高的OPR標記物計數趨於基線(紅線)。A點是異常標記物計數的高峰,且可以是“假陽性”點,如果病理和影像診斷沒有發現癌。B點是損傷癒合後的糾正點,且可以是“假陰性或低敏感”點,如果病理和影像診斷發現癌。對人群不同的取樣時間導致假陽性或假陰性。通過監測動態OPR標記物計數,可以看出體內的癌化狀態,且可以指導個體化治療指向正常基線至癌症痊癒。 4.4 移植瘤模型之謎 移植癌細胞在正常小鼠形成腫瘤並不容易。這被解釋為免疫抑制了腫瘤的形成[89]。然而,即使在免疫缺陷鼠,移植癌細胞也不一定很好地形成腫瘤[90, 91]。在某些情況下,移植瘤只在功能性T淋巴細胞存在下形成[92]。WOWH機制對此現象給出另一個解釋:正常鼠沒有正確的損傷分子環境使移植瘤反應,而裸鼠是部分損傷的身體,正好帶有合適的活化基因的某些癌細胞會對此有反應。由於是部分損傷的身體,裸鼠不提供適於所有移植癌細胞生長的分子,但提供損傷環境會促使瘤生長[92, 93]。在移植瘤模型上Matrigel的應用[94]即屬這種情況,因為Matrigel中的TGF-β[38], FGF[38], EGF[95], IGF-1[95],PDGF[96]和 NGF[97]在損傷中都積極表達,且它們都能激活癌基因,如Src[98–103],來啟動癌程序。EGF, FGF和PDGF的受體本身也是致癌基因(表-2)。由於Matrigel中有多種損傷相關細胞因子,一種或多種因子會符合移植瘤細胞先前活化的通路。因此,許多不成功的移植人細胞系在Matrigel存在下都易在鼠內生長[94],提示宿主的免疫缺陷與否不是腫瘤形成的限制因素,因為Matrigel更象是免疫刺激劑而不是免疫抑製劑[104–106]。此外,有實驗顯示損傷的個體(a)只需較少的移植細胞來形成腫瘤[68],和(b)比正常個體形成較大的腫瘤[92]。可以推測移植瘤細胞甚至可以在異原和免疫活性鼠上生長,只要有相應的損傷存在,尤其是慢性損傷。 5 總結 WOWH機制是一個基於從癌症,癌前病變,致癌基因,損傷修復和所有多細胞物種癌症發生的關係中推演而來的邏輯。損傷觸發了致癌基因細胞因子來徵募和分化幹細胞以癒合損傷。如果損傷癒合了,此過程會回復到自我平衡狀態。如果損傷持續,損傷和正反饋分子會使癌腫越來越大直到損傷癒合或整個系統衰竭。WOWH機制的邏輯與機體其它生理代謝機制一致—為了生存。它幫助理解許多突變理論所致的癌症謎團。它可以用於解釋癌症預防,復發,轉移,體外體內研究和個體化治療。
文獻請參閱英文原版: http://www.springerlink.com/content/lx671t71467n172k/fulltext.pdf
|


|
|
|
|
 |
 |
| 實用資訊 | |
|
|
|
|
| 一周點擊熱帖 | 更多>> |
|
|
|
| 一周回復熱帖 |
|
|
|
|
| 歷史上的今天:回復熱帖 |
| 2010: | 四個男性健康好習慣 | |
| 2010: | 重視養生又不刻意養生 | |
| 2009: | 金立鑫:我上《中醫概論》課的體會 | |
| 2009: | 杏林孤蟬:中醫是最大的忽悠 | |
| 2008: | 珍愛生命,遠離中醫藥 / 作者:古城 | |
| 2008: | 請有同樣經歷或熟知醫保條款的人士給個 | |
