接續:科學界為什麼,為誰捂蓋新冠病毒來源真相(上)
本文的評析對象是如下法廣(RFI)報道:
新冠溯源: 科學家們為何要壓制實驗室泄密理論?
https://www.rfi.fr/cn/%E5%9B%BD%E9%99%85/20220114-%E6%96%B0%E5%86%A0%E6%BA%AF%E6%BA%90-%E7%A7%91%E5%AD%A6%E5%AE%B6%E4%BB%AC%E4%B8%BA%E4%BD%95%E8%A6%81%E5%8E%8B%E5%88%B6%E5%AE%9E%E9%AA%8C%E5%AE%A4%E6%B3%84%E5%AF%86%E7%90%86%E8%AE%BA
該報道通篇“無數次”使用概念偷換、事實偷換等欺騙手法,反覆閹割、篡改事實,無中生有,一而再,再而三地避重就輕、轉移視線、栽贓嫁禍,以捂蓋新冠病毒來源的內在真相。下面繼續展示、剖析報道中的謊言、詭辯、欺詐,捂瞞、誤導、欲蓋彌彰。
謊言五
報道中說:
根據法拉爾的一份在2月2日星期日發給美國國家衛生研究院院長弗朗西斯-柯林斯和美國國家過敏和傳染病研究所所長安東尼-福奇的電子郵件的記載,杜蘭大學的羅伯特-加里(Robert Garry )和斯克里普斯研究所的邁克爾-法贊-的關注點集中在SARS-CoV-2基因組的一個特徵上,這個特徵以前從未在任何其他類似SARS的冠狀病毒中出現過:插入了一個12個字母的基因序列(與蝙蝠中最接近的病毒相比),創造了一個叫做furin酶切入點使得病毒的傳染性大大增強。Farzan認為,"在實驗室外很難解釋這個問題",Garry "想不出一個合理的自然情景......想不出這在自然界是如何完成的"。。。微生物學家Andrew Rambaut 也認為從微生物演變的角度來看,病毒酶切入點的出現是“十分不尋常的”!並且暗示只有武漢病毒學者才能夠作出解釋。
“微生物學家Andrew Rambaut 。。。暗示只有武漢病毒學者才能夠作出解釋”這句話,要麼是Andrew Rambaut罔顧事實、掩耳盜鈴的惡毒栽贓,要麼是RFI報道無中生有、憑空杜撰的下作誣衊。
能解釋新冠病毒中furin酶切位點的人,應該是做過furin酶切位點研究的人。武漢病毒研究所應該為新冠的furin酶切位點作出解釋嗎?武漢病毒研究所從未做過在冠狀病毒中插入furin酶切位點的研究、實驗,你叫他們怎麼解釋?真正能解釋,真正應該解釋新冠病毒中furin酶切位點來源的人,不是武漢病毒學者,而是美國病毒學家們。因為美國病毒學家們十多年來做過不計其數的在冠狀病毒或其它病毒中插入furin酶切位點的病毒改造研究、實驗,美國病毒學家對furin酶切位點的研究、實驗極為深入、全面,涵蓋以下幾種情況:
1、在沒有furin酶切位點的(冠狀)病毒中插入一個或多個furin酶切位點;
2、在已有furin酶切位點的(冠狀)病毒中插入新的furin酶切位點;
3、改造(冠狀)病毒中已有的furin酶切位點的酶切組合,對比改造前後不同furin酶切組合的水解、切割效率,致病能力、效果。。。
舉兩個例子。
第一個例子。2006 年3月,Montana大學的Jack Nunberg等人在Elsevier上發表了如下論文:
Furin cleavage of the SARS coronavirus spike glycoprotein enhances cell–cell fusion but does not affect virion entry
SARS病毒刺突(糖)蛋白的弗林切割增強了細胞-細胞融合,但不影響病毒進入
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7111780/
該研究在SARS-CoV刺突蛋白(Spike蛋白,S蛋白)S1/S2位置處(S蛋白S1、S2亞基交界處)插入了一個furin(弗林)酶切位點,並實驗證明,這一FCS(furin cleavage site,即furin酶切位點)插入顯著增強了SARS病毒誘導宿主細胞發生細胞–細胞融合,形成多核擴大細胞--合胞體的能力(細胞-細胞融合形成合胞體能提高病毒在宿主組織、器官中的擴散效率)。
新冠病毒的furin酶切位點也位於S蛋白S1、S2亞基交界處,與該論文的furin酶切位點插入位置完全相同。
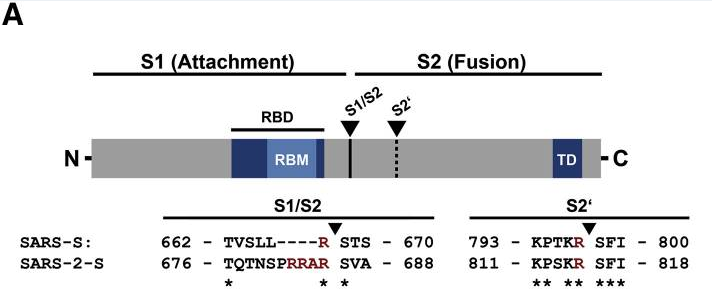
新冠病毒(SARS-2)S1/S2位置處的“RRAR”furin酶切位點
SARS病毒本身沒有S1/S2處的furin酶切位點,
與新冠同譜系、同支系(冠狀病毒β譜系B支系)的新冠近親病毒全都沒有S1/S2處的furin酶切位點;
與新冠同譜系(冠狀病毒β譜系)的蝙蝠冠狀病毒全都沒有S1/S2處的furin酶切位點;
與新冠刺突蛋白相似度>40%的新冠所有天然近親、遠親也全都沒有S1/S2處的furin酶切位點。

上圖是新冠病毒(Wuhan-Hu-1毒株)與進化樹中的“親戚”冠狀病毒在S1/S2附近氨基酸序列的對照圖,最上方的紅色箭頭指示的就是S1/S2位置(S1、S2亞基交界處)。由圖可見,在S1/S2處有furin酶切位點的唯有新冠病毒,圖中的新冠近親、遠親,無一在在S1/S2處有furin酶切位點(全都缺失furin酶切位點對應的氨基酸)。
請大家作個判斷,新冠病毒S1/S2處的furin酶切位點是怎麼得來的?是自然演化產生的,還是人為插入的?
第二個例子。2009年4月,Cornell大學的加里·惠特克(Gary Whittaker)等人在PNAS(美國國家科學院院刊)上發表了如下論文:
Activation of the SARS coronavirus spike protein via sequential proteolytic cleavage at two distinct sites
經由兩個不同位點的連續蛋白水解切割(觸發)的SARS 病毒刺突蛋白激活(活性增強)
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2660061/
該研究在SARS病毒刺突蛋白的兩個位置插入了furin酶切位點,一個在S1/S2處,另一個在S2亞基內(SARS刺突蛋白的第793~797氨基酸之間),並構造了三個SARS病毒的人工變異(改造)體,其中兩個各含一個furin酶切位點插入,另一個兼具兩個furin酶切位點插入。研究實驗證明:
1、S1/S2位置插入的furin酶切位點可使S蛋白被有效水解切割,導致S1、S2亞基相互分離;
2、S1/S2位置插入的furin酶切位點顯著增強了細胞-細胞融合,促進了合胞體的生成;
3、S1/S2位置插入的furin酶切位點顯著提高了SARS病毒的感染效率;
4、S2′位置插入的furin酶切位點可進一步增強細胞-細胞融合,促進合胞體生成,兼具兩個furin酶切位點的SARS改造體可導致超過95%的(被感染組織中的)細胞發生細胞-細胞融合,形成非常大的合胞體。
美國科學家所做的furin酶切位點相關的研究、實驗還有很多,我閱讀過的研究例證,將匯集、整理在稍後的其它文章中。
對新冠病毒S1/S2位置處,其近親無一具備的furin酶切位點,誰能夠作出解釋,誰應該作出解釋,作出澄清?是報道所說的“武漢病毒學者”,還是做過大量furin酶切位點插入、改造實驗的美國病毒學家們?
謊言六
報道中說:
從最新公布的郵件上可以看出,美國國家過敏和傳染病研究所所長安東尼-福奇從第一時間便警告專家不應該公開討論病毒來自實驗室的理論,因為這會對科學研究,尤其是中國的科學研究造成攻擊。這也是為何福奇本人曾經十多次強調病毒只能是自然來源,而美國國家衛生研究院院長弗朗西斯-柯林斯更是將實驗室泄露理論定性為荒謬。
“自然來源論”的反面是“實驗室來源論”,不是“實驗室泄漏論”。由艾滋病、西班牙流感的先例可知,病毒的最早發現地、最早暴露地不等於病毒的來源地、產生地(設計、製造地)。
如果新冠病毒不是自然產生的,那麼,它就是實驗室產生的;如果新冠病毒是實驗室產生的,那麼,它可能來自武漢的某個實驗室,它同樣可能來自武漢之外、中國之外的某個實驗室。
沒有任何證據,沒有任何理由可排除新冠病毒來自武漢之外實驗室的可能性。新冠病毒來自武漢之外實驗室的可能性,是新冠溯源工作應深入調查,不可忽略、不容遺漏的必要內容。
“實驗室泄漏論”只是“實驗室來源論”的一個分支。來自實驗室,不等於來自“實驗室泄漏”。“實驗室泄漏”,相當於“本地實驗室泄漏”,“實驗室泄漏論”的潛台詞是:新冠病毒來自武漢的某座實驗室。用“實驗室泄漏論”偷換“實驗室來源論”,將二者混為一談,是將“實驗室來源論”蘊含的多種可能性閹割為“武漢實驗室來源”單一可能性,是在隱蔽、捂蓋新冠病毒的其它實驗室來源可能性。
在以往的文章中,我曾從不同角度,以不同方式,基於不同證據,多次論證過以下兩個結論(最近的一次論證是在上篇文章的“謊言一”小節給出的):
1、新冠病毒不是武漢病毒研究所設計、製造的;
2、新冠病毒不可能是武漢病毒研究所泄漏的;
這是兩個得到大量堅實證據交叉支撐,經得起反覆推敲的結論。由這兩個結論可知,“實驗室泄漏論”根本不成立!
美國及美國的盟友們為什麼無休止地炒作“實驗室泄漏論”?他們為什麼對否定“實驗室泄漏論”的眾多鐵證視而不見?為什麼美國政府一直抱怨缺乏足夠的溯源證據、資料?因為美國溯源調查的目的不是揭示、還原新冠病毒來源真相,新冠疫情產生真相;美國需要的是製造替罪羊的證據,不會引火燒身的證據,所有不利於美國,不利於美國製造替罪羊,不利於美國甩鍋嫁禍的證據,統統不是證據。
新冠病毒來自實驗室,它又不是武漢病毒研究所設計、製造、泄漏的。那麼,新冠病毒肯定來自武漢病毒研究所之外的實驗室,它極有可能來自武漢之外、中國之外的實驗室!
弗朗西斯-柯林斯(Francis Collins)力挺的是“自然來源論”,他曾在2020年3月26日發表名為《基因研究顯示 新冠肺炎病毒起源於自然》的博客文章,宣稱:該病毒是自然產生的。
美國國立衛生研究院院長柯林斯:新冠肺炎病毒源於自然_新聞中心_中國網
http://news.china.com.cn/2020-03/28/content_75871483.htm
對弗朗西斯-柯林斯來說,正論是“自然來源論”,謬論是“實驗室來源論”。RFI報道再一次用“實驗室來源論”的分支“實驗室泄漏論”以偏蓋全、偷換“實驗室來源論”,將新冠病毒的來源與武漢病毒研究所單一掛鈎,籍此隱匿、掩藏新冠病毒的其它實驗室來源可能性。
壓制“實驗室泄漏論”,是為武漢病毒研究所打掩護,是為中國打掩護;壓制“實驗室來源論”,是為從事冠狀病毒功能增益改造研究的實驗室打掩護,是為開展舉國性質危險病原體功能增益改造研究,以聯邦資金資助本國功能增益改造研究的國家打掩護。
弗朗西斯-柯林斯、安東尼-福奇全力壓制的,是“實驗室泄漏論”,還是“實驗室來源論”呢?他們是在為武漢病毒研究所、為中國打掩護,還是在為美國實驗室,為美國打掩護呢?
報道中說,安東尼-福奇第一時間警告專家不應該公開討論病毒來自實驗室的理論,福奇本人曾經十多次強調病毒只能是自然來源,美國國家衛生研究院院長弗朗西斯-柯林斯更是將實驗室泄露理論定性為荒謬。二人如此積極、如此投入,如此不遺餘力地壓制、捂蓋、撒謊,不惜身敗名裂也要欺騙全世界,他們真的是為保護中國的科學研究嗎?他們真的是為武漢病毒研究所打掩護嗎?他們可能為中國如此捂蓋新冠病毒的來源真相嗎?這合乎情理嗎?這說得通嗎?
他們是忘我奉獻、舍已為人的“活雷鋒”,為中國兩肋插刀、至死不渝的“白求恩”嗎?
謊言七
報道中說:
2020年3月17日自然雜誌發表的標題為“The proximal origin of SARS-CoV-2”的文章對病毒來源問題一錘定音,文章雖然也對病毒基因序列組成與furin酶切入點提出疑問,但認為可以在自然演變中完成。
實驗室泄露的可能性被徹底排除在外,文章的簽名者中就包括微生物學家Andrew Rambaut 與病毒學家羅伯特-加里(Robert Garry )等對病毒持有極大懷疑的專家,究竟是何等原因使他們在一個月內改變看法呢?羅伯特-加里日前向美國攔截組織The Intercept.表示他當初對酶切入點的印象使(是)錯誤的,他說科學就是如此逐漸走向進步的,無人故意要愚弄公眾,他今天依然認為他們在自然雜誌上發表的文章是有關新冠病毒的最完美的分析文章。
(報道引用結束)
Nature論文“The proximal origin of SARS-CoV-2(SARS-CoV-2的近端起源)”地址如下:
https://www.nature.com/articles/s41591-020-0820-9
中文介紹可參考:
Nat Med:研究證實SARS-CoV-2屬自然進化,不可能是人為製造! - 組學專區 – 生物谷
https://news.bioon.com/article/6752206.html
美英澳科學家《自然醫學》再添力證:新冠病毒乃自然進化產物…… 廣東省科學技術廳
http://gdstc.gd.gov.cn/kjfyzl/gnwdt/content/post_2936176.html
論文力撐自然來源論,宣稱:對SARS-CoV-2及相關病毒的基因組序列進行分析後發現,沒有證據表明該病毒是實驗室製造或實驗室(方式)產生的。
顯而易見,論文否定、排除的不是“實驗室泄露論”,而是“實驗室來源論”。RFI報道第N次使用概念偷換、事實偷換手法,以偏代全地謊稱“實驗室泄露的可能性被徹底排除在外”。
這麼一偷換,一篇捂蓋新冠“實驗室來源”,為美國危險病原體功能增益改造研究打掩護的論文,搖身一變,變成了捂蓋“武漢實驗室泄漏”,為武漢病毒研究所打掩護的論文;論文的包庇、服務對象,由開展功能增益研究的美國實驗室,變成了沒有做過功能增益研究,沒有發表過任何功能增益研究論文的武漢病毒研究所;論文的包庇、服務國,從開展舉國性質功能增益研究的第一生物科技強國--美國,變成了功能增益研究屈指可數、生物科技水平落後美國十幾年的中國。經偷換、篡改捏造出的包庇、服務對象作為新冠病毒來源的唯一嫌疑犯被RFI押上了被告席,而論文真正的包庇、服務對象則悄然隱身於世人目光之外。
論文的作者是以下五人:美國斯克利普斯研究所的克里斯蒂安·安德森(Kristian G Andersen)、英國愛丁堡大學進化生物學研究所的Andrew Rambaut、美國哥倫比亞大學公共衛生學院感染與免疫中心主任W.伊恩·利普金(W. Ian Lipkin)、澳大利亞悉尼大學的愛德華·霍姆斯(Edward C. Holmes),美國杜蘭大學醫學院微生物學和免疫學系的羅伯特·f·加里(Robert F. Garry)。
五人中三人是美國病毒學家,另兩人分屬英國、澳大利亞。“巧合”的是,五人的所屬國澳大利亞(AU)、英國(UK)、美國(US)恰好是AUKUS軍事聯盟(澳英美軍事聯盟)的三個成員國。AUKUS軍事聯盟的首要戰略敵人是中國,美、澳、英三國是近幾年打擊中國現政府最不遺餘力的三個國家。試想,這三個國家的病毒學家可能聯手為武漢病毒研究所打掩護,聯手為中國捂蓋新冠病毒的來源真相嗎?
他們究竟在為誰捂蓋?
五位論文作者中,當初認為新冠病毒來自實驗室的至少有三位。上述報道文字提到了兩位:愛丁堡大學的Andrew-Rambaut,杜蘭大學的羅伯特·加里(Robert Garry)。還有一位是上篇文章提到過的,曾在2020年1月31日致信安東尼-福奇,表示新冠病毒有基因改造特徵,不符合進化理論,但在次日與福奇電話會議後迅速改變立場的斯克利普斯研究所的克里斯蒂安·安德森(Kristian G Andersen)。
甘當政客們的科學工具,可以隨時出賣真相、事實、是非的三個牆頭草有科學操守和基本人格可言嗎?
報道中提到,羅伯特-加里說自己並非“故意要愚弄公眾”,並聲稱“他今天依然認為他們在自然雜誌上發表的文章是有關新冠病毒的最完美的分析文章。”
如此“完美”的論文,不近距離欣賞、領略一番將是巨大的遺憾。羅伯特-加里等五人的Natue大作到底有多完美呢?它至少有三大“完美表現”。
完美表現一
論文稱,新冠病毒的骨架與已知病毒的骨架都有很大不同,因此,它不是基於某種已知病毒骨架構建的(進而它不是實驗室構建的)。
論文隱含了一個假設:實驗室設計、製造不出與已知病毒的骨架都有很大不同的病毒。這一假設的推論是:與已知病毒的骨架都有很大不同的病毒一定不是實驗室構造的。
論文使用了上述假設的推論來否定新冠病毒來自實驗室。問題是,上述假設是錯誤的。
實驗室能不能設計、製造出與已知病毒的骨架都有很大不同的病毒呢?太能了!對現代基因改造技術來說,這根本就不是個問題。我舉個例子。
2008年12月16日,Ralph S. Baric(冠狀病毒研究全球第一人,冠狀病毒功能增益改造狂人)領導的團隊發表了一篇PNAS(美國國家科學院院刊)論文:
Synthetic recombinant bat SARS-like coronavirus is infectious in cultured cells and in mice
合成的重組蝙蝠類SARS冠狀病毒在培養細胞和小鼠中具有傳染性。
https://www.pnas.org/content/105/50/19944
論文通過兩步改造,合成了一個重組病毒Bat-SRBD。Bat-SRBD是用5種病毒原材料重組合成的,它們是SARS-CoV和4種蝙蝠冠狀病毒:HKU3-1、HKU3-2、HKU3-3和RP3 。重組病毒Bat-SRBD的RBD來自SARS病毒,骨架(其它部分)來自4種蝙蝠冠狀病毒。
實驗證明,Bat-SRBD能有效感染HAE細胞(人氣道上皮細胞)中的纖毛細胞(但不能感染同屬HAE細胞的非纖毛細胞)、非洲綠猴的Vero E6細胞(一種腎細胞)、表達人類ACE2的轉基因小鼠DBT細胞(延遲腦腫瘤細胞)DBT-hACE2、表達果子狸ACE2的轉基因小鼠DBT細胞DBT-cACE2,並在以上各細胞組織中大量複製。
也就是說,重組病毒Bat-SRBD是一種能感染人類、非洲綠猴、果子狸(civet)三種動物,複製能力強大,可跨物種感染、傳播的實驗室改造病毒。
由這篇論文可知,改造病毒不必只基於一種病毒(骨架),可以基於多種病毒來獲得改造病毒,改造病毒將兼具多種病毒的結構,改造病毒的結構將與素材病毒中的任何一種都有明顯差異。早在2008年,病毒學家就可以很容易地設計、製造出混成式的冠狀病毒了。而且,Ralph S. Baric當時就有能力設計、製造出比Bat-SRBD複雜100倍的病毒。2000年發明基於基因序列合成冠狀病毒的反向遺傳平台之後,Ralph S. Baric等人已將冠狀病毒的設計、改造簡化為基因序列文本文件的設計、改造了。
因此,論文的假設是錯誤的,論文作者是在故意裝糊塗。為了顛倒黑白,“論證”自然來源論,裝糊塗是必須的。
新冠病毒的骨架與已知病毒的骨架都有很大不同,原因與上述重組病毒Bat-SRBD類似。我之前的文章多次指出,新冠病毒是跨科屬的病毒集大成者,它借鑑、引入、集成了多種冠狀病毒、多種非冠狀病毒的結構和致病特性,它不是一個基於單一原材料骨架的簡單改造體。存在多重結構來源(結構來源五花八門),部分結構甚至借鑑、來自非冠狀病毒(包括艾滋病病毒、埃博拉病毒等等)的新冠病毒的骨架當然與已知病毒的骨架都有很大不同。
實際上,“新冠病毒的骨架與已知病毒的骨架都有很大不同”,這一事實不是新冠病毒自然產生的證據,而是新冠病毒實驗室產生的證據。因為自然演化產生的病毒必然存在高度相像的祖先病毒、近親病毒,只有在實驗室經過多重改造(多重集成、多重嵌合、多重編輯)得到的病毒才會和所有自然病毒無一高度相像。
完美表現二
論文稱,新冠病毒的RBD(Receptor Binding Domain,受體結合域,決定S蛋白即刺突蛋白ACE2結合能力的S1亞基的一部分)不是最優的,如果新冠病毒是人為設計的,那麼,它的RBD應該設計得更好。
論文此處的邏輯(又一個隱含的假設)非常“完美”、“嚴謹”:人為設計的病毒必然是最優的;不是最優設計的病毒一定不是人為設計的。
按照這一邏輯,可得出如下推論:實驗室設計、製造的病毒,只有終極版本才(可能)是實驗室設計、製造的,其它版本都肯定不是實驗室設計、製造的,肯定都是自然產生的。因為非終極版本的設計都不是最優的。
新冠病毒的RBD是不是真的不夠好呢?事實上,新冠的RBD設計即使不是最優的,也是極為接近最優的,這體現在:
1、新冠刺突蛋白與人類ACE2的結合親和力(結合親和力由刺突蛋白的RBD決定),是SARS刺突蛋白與人類ACE2結合親和力的10~20倍;
2、新冠刺突蛋白與人類ACE2的結合親和力,強於它與常見動物ACE2的結合親和力,人類ACE2是新冠刺突蛋白的最佳結合對象,人體細胞是新冠病毒的最佳感染對象;
3、新冠病毒一經出現,未經過任何宿主跳躍所需的突變,其刺突蛋白便極為適合結合人類ACE2,新冠病毒便極為適合感染人類。
(由2、3可得出如下推論:新冠病毒的原始宿主很可能就是人類,新冠病毒很可能是專為感染人類而設計的。)
第1點的依據是,2020年3月13日,美國德克薩斯大學奧斯汀分校Jason S McLellan團隊,與美國國家過敏和傳染病研究所的研究人員在《Science》雜誌上聯合發表的如下論文:
Cryo-EM Structure of the 2019-nCoV Spike in the Prefusion Conformation
(膜融合前構象中2019-nCoV 刺突蛋白的冷凍電鏡結構)
https://science.sciencemag.org/content/367/6483/1260
第2、3兩點的依據是,澳大利亞弗林德斯大學、拉籌伯大學等機構的科學家2021年6月24日發表的如下Nature雜誌論文(該論文早在2020年5月13日就已發表於預印本平台arxiv):
In silico comparison of SARS-CoV-2 spike protein-ACE2 binding affinities across species and implications for viral origin
(新冠刺突蛋白與ACE2結合親和力的跨物種模擬比較及其對病毒起源的指示)
https://www.nature.com/articles/s41598-021-92388-5
完美表現三
論文指出,新冠病毒S1、S2亞基交界處furin酶切位點周圍存在一個具有免疫屏蔽功能(可屏蔽宿主免疫系統搜索、發現、攻擊)的O-Linked聚糖結構。論文稱,在實驗室環境中將祖本病毒置於培養細胞中連續傳代不太可能變異產生這一結構,這一具有免疫屏蔽功能的結構需要在(在人體活體)免疫系統的參與下才能變異產生。因此,新冠病毒不是實驗室培養產生的。
論文作者又裝糊塗了,他們使用了第三個不成立的假設,把實驗室培養(以一定方式、壓力培養病毒,促使其在連續傳代過程中發生變異)作為獲得實驗室病毒,及人為產生O-Linked聚糖結構的唯一途徑。事實上,這種正向遺傳學方式早就不是病毒改造的主流手段了(有時仍作為輔助手段),它早已被直接編輯基因組或編輯基因序列的反向遺傳技術取而代之了;病毒學家們根本不會,根本無須用這種難以達到既定目的的方式去產生O-Linked聚糖結構,他們可以快捷地、輕而易舉地在病毒中直接構造出O-Linked聚糖結構。
我們近距離觀察一下新冠病毒刺突蛋白(S蛋白)中的O-Linked聚糖結構,這一結構由furin酶切位點(RRAR)兩側的三個氨基酸構成:S蛋白氨基酸序列的第673氨基酸-絲氨酸(S),第678氨基酸-蘇氨酸(T),第687氨基酸-絲氨酸(S)。這三個氨基酸在下圖中用紫色方框標出:

新冠furin酶切位點周圍的一個蘇氨酸(T),二個絲氨酸(S)形成一個O-Linked聚糖結構
新冠刺突蛋白的第678氨基酸是蘇氨酸(T),而SARS和絕大多數類SARS冠狀病毒(新冠和SARS同處β譜系B支系,這一支系的病毒統稱類SARS冠狀病毒)對應位置處的氨基酸是絲氨酸(S);新冠刺突蛋白的第673、687氨基酸則和SARS等近親相同,都是絲氨酸(S)。
註:上圖中另二個新冠近親Bat-RaTG13和Pangalin(代表一種馬來穿山甲冠狀病毒)的第678氨基酸也是蘇氨酸(T),與新冠相同,但這兩個病毒沒有病毒毒株,而且它們的基因序列是疫情發生後才宣布的,是專門用來證明新冠“自然來源論”的,它們的真實性非常可疑。
基於新冠的近親病毒(如SARS或蝙蝠冠狀病毒)構造O-Linked聚糖結構,只要把對應新冠病毒刺突蛋白第678氨基酸的近親病毒氨基酸由絲氨酸(S)替換為蘇氨酸(T)就可以了(忽略O-Linked聚糖結構與furin酶切位點的關聯性)。至少有兩種反向遺傳技術可快速實現這一替換:
一是使用基因剪刀(CRISPR-Cas9 )直接編輯近親病毒基因組,完成氨基酸替換;
二是先編輯近親病毒的基因序列,在基因序列(文本)中設計出這一結構(把絲氨酸的核苷酸密碼子符號替換成蘇氨酸的密碼子符號就ok了),再基於設計好的新的基因序列使用反向遺傳平台合成病毒基因組、收穫病毒毒株,O-Linked聚糖結構就在所收穫的病毒毒株中存在了。
HIV-1病毒(即艾滋病病毒,屬於逆轉錄病毒科、慢病毒屬)、埃博拉病毒(絲狀病毒科病毒)、丙型肝炎病毒(黃病毒科)等非冠狀病毒也有O-Linked聚糖結構及免疫屏蔽能力,新冠的O-Linked聚糖結構很可能借鑑自這些非冠狀病毒。
五位病毒大腕的Nature論文不是學術作品,不是寫給同行看的。它是某國領導下的全球科學界的“大外宣”工具,它的存在價值是拉大旗作虎皮,作為“權威著作”供“自然來源論”反覆引用, 為“自然來源論”撐腰、壯膽、開道。
(未完待續)






