最近的一項研究向世人展示了新冠病毒來自實驗室的又一鐵證。
2022年2月21日,來自美國、印度、瑞士、意大利四國的多所大學、醫療機構的一組科學家、學者在frontiers in Virology(病毒學前沿)期刊發表了如下論文:
MSH3 Homology and Potential Recombination Link to SARS-CoV-2 Furin Cleavage Site
MSH3同源性及與SARS-CoV-2弗林酶切割位點的潛在重組聯繫
https://www.frontiersin.org/articles/10.3389/fviro.2022.834808/full
因發現、揭示了新冠基因序列與莫德納專利序列的重大關聯,這篇論文如今已廣為人知。
論文核心要點是:
BLAST(核酸序列、蛋白序列搜索、比對工具)搜索顯示,SARS-CoV-2基因組中包含furin切割位點在內的19個核苷酸,與莫德納專利中一段序列的反向序列完全(100%)互補匹配。
怎麼理解這段話呢?我們先來看看論文所說的新冠病毒基因組中的19個核苷酸。
新冠病毒(SARS-CoV-2)基因組S1/S2處(S蛋白S1、S2亞基交界處)存在如下19個核苷酸:CU CCU CGG CGG GCA CGU AG,其中標記為黑體的12個核苷酸(CCU CGG CGG GCA)對應新冠病毒S1/S2處的獨特四氨基酸插入“PRRA”(脯氨酸-精氨酸-精氨酸-丙氨酸)。“PRRA”插入的後三個氨基酸“RRA”,與基因序列中其後相鄰的“R”(精氨酸,對應19個核苷酸中的CGU),構成了新冠病毒的furin酶切組合“RRAR”,即新冠S1/S2處的furin酶切位點。furin酶切組合的模式是“RXXR”,它要求四氨基酸組合的首尾氨基酸都是精氨酸(R)。
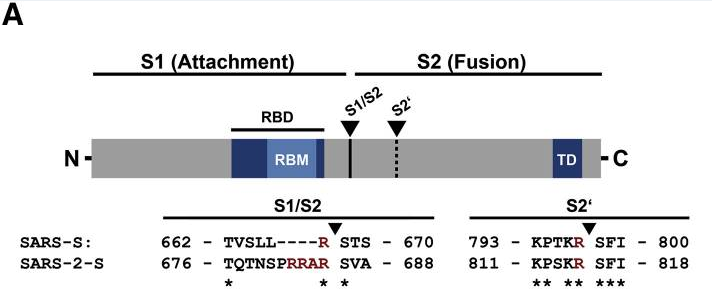
新冠病毒S1/S2處的“PRRA”插入及“RRAR”furin酶切位點
為什麼12個核苷酸對應四個氨基酸呢?因為一個氨基酸(殘基)對應三個核苷酸的組合,如CGG對應精氨基酸(R),表達氨基酸的三核苷酸組合稱為氨基酸的密碼子;同一個氨基酸可以用不同的密碼子表達,例如,以下六種密碼子都可以表達精氨基酸(R):CGU,CGC,CGA,CGG,AGA和AGG。新冠“RRAR”furin酶切組合對應前面19個核苷酸中的如下12個核苷酸CGG CGG GCA CGU,其中前兩個精氨酸(R)是用CGG密碼子表達的,最後一個精氨酸則是用CGU密碼子表達的。
接下來,我們看看莫德納專利與新冠病毒基因組有什麼關聯。
研究者發現,新冠病毒S1/S2處的上述19個核苷酸竟然在2016年莫德納公司申請的一項如下美國專利(patent US 9587003,Filed: February 4, 2016)中(互補)出現過:
Modified polynucleotides for the production of oncology-related proteins and peptides
用於生產腫瘤相關蛋白質和多肽的修飾多核苷酸
https://uspto.report/patent/grant/9587003
https://patents.google.com/patent/US9587003B2/en
在專利US 9587003 中,有一個包含約3300個核苷酸的DNA核苷酸序列Seq ID11652,這個序列的第2733至2751核苷酸是如下19個核苷酸:CT ACG TGC CCG CCG AGG AG,這19個核苷酸的短序列倒過來寫是:GA GGA GCC GCC CGT GCA TC,後者正好是新冠病毒S1/S2處19個核苷酸的互補序列,即後者正好與新冠中的那19個核苷酸完全互補匹配。
為方便比較觀察,將所涉及的三組19個核苷酸序列對齊展示如下:

新冠病毒是單鏈RNA病毒,其基因序列是RNA核苷酸序列,莫德納專利中的序列是DNA核苷酸序列。RNA鏈或RNA序列與DNA鏈或DNA序列的匹配不是鹼基(鹼基是核苷酸的核心構件,鹼基、核苷酸這兩個術語常常相互代表,不特別區分)等同匹配,而是鹼基互補匹配(與RNA對應的DNA叫做cDNA,即Complementary DNA,RNA的互補DNA)。具體來說,RNA鏈或序列中的鹼基與其互補DNA鏈或序列中鹼基的匹配關係是:
A-T(即RNA中的腺嘌呤A匹配DNA中的胸腺嘧啶T)、U-A、G-C、C-G。
代表鹼基(及以鹼基為核心的核苷酸)的各字母含義如下:腺嘌呤(A)、胸腺嘧啶(T)、鳥嘌呤(G)、胞嘧啶(C)、尿嘧啶(U)。
胸腺嘧啶(T)是尿嘧啶(U)5號位碳原子甲基化的衍生物。胸腺嘧啶(T)基本上只出現在DNA基因中(在RNA基因中極罕見),尿嘧啶(U)基本上只出現在RNA基因中,但在序列表達時,RNA(核糖核酸)的尿嘧啶符號U通常不使用,統一以DNA(脫氧核糖核酸)中胸腺嘧啶的符號T代表(代替)。
通過對比不難發現,上圖中的前二個序列,即新冠病毒S1/S2處19個核苷酸的短序列(RNA序列)CU CCU CGG CGG GCA CGU AG,與莫德納專利中19個核苷酸的短序列(DNA序列)的反向序列GA GGA GCC GCC CGT GCA TC正好是完全互補匹配的,這兩個序列的所有對應核苷酸(鹼基)都符合A-T、U-A、G-C、C-G的匹配關係。
下圖是論文附圖,它展示了新冠S1/S2處的aa(amino acids,氨基酸)序列、核苷酸序列,及莫德納專利(patent US 9587003)中相應的19個核苷酸的短序列的反向序列(已轉化為互補序列)。另外,圖中新冠核苷酸序列中的尿嘧啶U已被胸腺嘧啶符號T所代替(代表),新冠S1/S2處的19個核苷酸CU CCU CGG CGG GCA CGU AG在圖中表示為:CT CCT CGG CGG GCA CGT AG。

論文指出,莫德納專利中包含“19個核苷酸”的序列SEQ ID11652與MSH3蛋白的核苷酸序列有關,該序列似乎是MSH3序列針對人類做過密碼子優化後得到的(這意味着SEQ ID11652序列不是天然生物序列)。MSH3是mutS homolog 3的縮寫,是DNA錯配修復蛋白MutS 的人類同源物,它的作用是糾正DNA合成過程中的鹼基錯配和缺失,許多腫瘤、癌症的發生與該蛋白的基因突變或功能故障有關。
莫德納專利中做過密碼子人類優化的MSH3序列(即SEQ ID11652)中的19個核苷酸的短序列的反向序列為什麼會(互補)出現在新冠病毒基因組中,而且恰恰出現在S蛋白S1、S2亞基交界處這一特殊位置,使新冠病毒的致病能力得到質的提升?這正常嗎?這合乎邏輯嗎?
論文作了專門的估算,估算結果是,莫德納專利中那19個核苷酸自然(互補)出現在基因組長度與新冠病毒相同(約30,000個核苷酸)的病毒基因組中的概率是:3.21 ×(10的負11次方)。
10的負11次方即千億分之一,也就是說,莫德納專利中的那19個核苷酸(的短序列)自然出現在新冠病毒基因組中的概率約為千億分之3.21。
註:粗略看了一下論文的算法,它似乎並未考慮19個核苷酸的出現位置,如果計算19個核苷酸出現在新冠特定位置(S1/S2處)的概率,那麼結果將是千億分之3.21×(1/30000)。
論文所指出的更關鍵的一點是:
新冠S1/S2處19個核苷酸的短序列(或其互補序列)在任何真核生物或任何其它病毒基因組中都不存在。
新冠不僅是擁有這一短序列的獨一無二的一種病毒,而且這一短序列還恰好出現在新冠S蛋白(刺突蛋白)S1、S2亞基交界處,這一對新冠致病能力至關重要的特殊位置。如我之前文章所述(如“新冠病毒來自何方”一文),新冠病毒基因結構中還有其它多個類似的“巧合”,這些巧合中的任何一個都難以甚至無法用非人為來解釋,然而,這些極低概率的“巧合”卻在新冠病毒中同時發生了。
新冠病毒來自何方
真核生物是具有(被膜包裹着的)細胞核的單細胞生物和多細胞生物的總稱,它包括所有動物、植物、真菌和其他具有由膜包裹着的(細胞核的)複雜亞細胞結構的生物,但不包括細菌和古菌,因為它們的細胞器(如細胞核)無膜包裹(摘自維基百科“真核生物”條目,略有修改)。
新冠中19個核苷酸的短序列是否在非真核生物中存在?我用NCBI Blast搜索了一下,發現在一些細菌(或古菌)DNA中確實存在19核苷酸短序列CT CCT CGG CGG GCA CGT AG,含有這一短序列的部分細菌(或古菌)的例子將在文尾提供。
細菌的遺傳物質是DNA,DNA是兩條鏈的雙螺旋,兩條DNA鏈彼此也是鹼基互補配對的,一條DNA鏈含有新冠中19核苷酸的短序列CT CCT CGG CGG GCA CGT AG,那麼另一條DNA鏈將含有莫德納專利中19核苷酸短序列的反向序列GA GGA GCC GCC CGT GCA TC。
回歸論文。論文指出,新冠病毒中19個核苷酸的短序列不可能是通過自然重組得到的(既不能重組自其它病毒,也不能重組自其宿主)。另外,論文雖未指出,但不難判斷,19個核苷酸的短序列也不可能繼承自新冠病毒假設的自然祖先,因為沒有任何其它病毒包含這一核苷酸短序列。
結論已經昭然若揭:
新冠病毒S1/S2處19個核苷酸的短序列(其中12個核苷酸對應新冠S1/S2處的furin酶切位點“RRAR”)不是自然產生、自然獲得的,它是人為引入的。
我之前的系列文章“誰設計、製造了新冠病毒(二)”指出,新冠病毒S1/S2處的“PRRA”插入及“RRAR”furin酶切位點是人為設計、引入的,下面重述一下相關要點,以與論文內容相互參照、印證:
一、新冠病毒的近親病毒全都沒有S1/S2處的furin酶切位點,具有S1/S2處furin酶切位點的冠狀病毒全都與新冠親緣關係極遠、差異巨大。具體而言:
1、與新冠同譜系、同支系(冠狀病毒β譜系B支系)的所有病毒除新冠外,全都沒有S1/S2處的furin酶切位點;
2、與新冠同譜系(冠狀病毒β譜系)的蝙蝠冠狀病毒全都沒有S1/S2處的furin酶切位點;
3、S蛋白與新冠S蛋白相似度>40%的所有冠狀病毒全都沒有S1/S2處的furin酶切位點。
二、新冠病毒S1/S2處的furin酶切位點不是自然演化(自然變異或自然重組)產生的,它是人為設計、引入的;而且,新冠所使用的“RRAR”或者說“PRRAR”酶切組合是一個經過精心遴選、精心設計的性能非常優異的furin酶切組合。
三、新冠病毒S1/S2處的“RRAR”酶切組合借鑑自小鼠肝炎病毒MHV-JHM株系、MHV-3株系,以及貓傳染性腹膜炎病毒FIPV(Feline infectious peritonitis virus)的某些病毒株,這三種(類)毒性極強、致死率極高的冠狀病毒S蛋白S1/S2處具有同樣的複合furin酶切組合“RRARR”,該組合的前四個氨基酸“RRAR”,被新冠病毒設計者引入、應用到了新冠病毒中,成為新冠S1/S2處的furin酶切組合(酶切位點)。
誰設計、製造了新冠病毒(二 下)
基於論文,我們可以得出一個新的要點:
四、新冠病毒S1/S2處的“PRRAR”氨基酸組合的密碼子表達(即氨基酸序列對應的核苷酸序列)CCU CGG CGG GCA CGU(如用胸腺嘧啶符號T代替尿嘧啶U則為:CCT CGG CGG GCA CGT),借鑑自莫德納專利US 9587003中一段短序列的反向序列(如前所述,該核苷酸短序列自然出現在新冠S1/S2處,非人為引入的概率極低)。專利中包含這段短序列的SEQ ID11652序列不是天然生物序列,它是針對人類做過密碼子優化的MSH3序列。
由上述論文能得出“莫德納公司製造了新冠病毒”這一結論嗎?我認為不能。莫德納專利是公開的,他人完全有可能專門研究該專利並借鑑、復用專利中的核苷酸序列片斷、反向序列片斷及互補序列片斷,即使該專利特別保護了19個核苷酸的短序列,借鑑、復用者也至多涉及付專利使用費的問題(我不清楚專利是否會保護專利中序列的反向序列)。更何況,新冠病毒是非公開的研究產物(新冠病毒應是被美國秘密盟友的海外科技間諜盜出的半成品),對新冠病毒設計者來說,將莫德納專利中核苷酸序列片斷的反向序列嘗試應用於新冠病毒,這並無為難之處。
那麼,新冠病毒究竟是誰設計、製造的?如我的文章“無數次”指出的,新冠病毒的核心設計、製造者,是論文內容、項目內容與新冠多項特殊結構、特性密切相關的“冠狀病毒合成之父”、功能增益改造狂人、現美國國家科學院院士拉爾夫·巴里克(Ralph S. Baric)。當然,我不能排除拉爾夫·巴里克與莫德納公司有合作關係及合作研發新冠病毒的可能性。
最後,我們來甄別一下“有據核查”對論文的解讀(頭一次聽說這個“有據核查”)。其解讀參見以下文章中的“延伸閱讀:有據核查:研究證實美國莫德納製造新冠病毒?真的嗎?”部分。
“研究證新冠是美制”登微博熱搜榜首 網民熱議 – 萬維讀者網
https://news.creaders.net/china/2022/03/24/2465381.html
“有據核查”稱:
不過,使用NCBI的BLAST 工具,Healthfeedback的專家在其他生物中找到了與Moderna專利的改良MSH3 基因高度相似的基因:

請大家注意,上圖列出的是與專利中改良MSH3(做過密碼子優化的MSH3)序列,即Seq ID11652序列(包含約3300個核苷酸)有一定匹配度的序列,不是與Seq ID11652序列中的19個核苷酸的短序列有一定匹配度的序列,更不是包含這19個核苷酸的序列。
而且,“有據核查”居然大言不慚地把71%~76%的較低匹配一致性(相似度)稱為“高度相似”。
“有據核查”在文章中並未展示任何包含“19個核苷酸”的生物序列。
而後,“有據核查”又說:
針對這篇論文,生物學專業科普公眾號“飛雪的博物大雜燴”總結:
1、無論是BLAST結果,還是已有的生物學研究都表明,FCS廣泛存在於生物界中,並且新冠病毒中的19nt序列(核心12nt)序列同樣存在於生物界中。
2、。。。
這段文字有兩個問題。
問題一:“FCS(furin cleavage site,furin酶切位點)廣泛存在於生物界中”。確實有不少病毒具有furin酶切位點,確實有不少冠狀病毒S1/S2處存在furin酶切位點;但是,如前文所述,新冠病毒的近親病毒無一具有S1/S2處的furin酶切位點,惟獨新冠具有S1/S2處的furin酶切位點,具有S1/S2處furin酶切位點的冠狀病毒全都與新冠親緣關係極遠、差異巨大,這是極不正常的。一對白人夫婦生出了一個黑皮膚小孩,這能用“黑人廣泛存在”解釋嗎?這是正常合理的事情嗎?這應當安之若素、不予深究嗎?
問題二:“並且新冠病毒中的19nt序列(核心12nt)序列同樣存在於生物界中。“有據核查”前面搜索、列出的是一些與密碼子優化過的MSH3 序列,即Seq ID11652序列相似度約為71%~76%的序列,並非包含“新冠病毒中的19nt”的序列(nt即nucleotide,核苷酸),“有據核查”根本未查出、展示任何“包含新冠病毒中19個核苷酸”的生物序列,它也展示不出包含這19個核苷酸的真核生物序列或病毒序列。
如前所述,論文明確指出:
沒有在任何真核生物或任何新冠以外的其它病毒基因組中發現新冠中的19個核苷酸的短序列(或其互補序列)。
儘管在某些細菌或古菌(二者是非真核生物)中存在相同的19核苷酸短序列,但是,沒有任何其它病毒,沒有任何真核生物具有這一短序列。試問:
新冠病毒是從哪一祖先病毒那裡繼承獲得這一短序列的?
新冠假設的祖先是如何通過重組從其它病毒那裡獲得這一短序列的?
新冠假設的祖先是如何從其動物宿主(真核生物)那裡以重組或其它方式獲得這一短序列的?
如果這一短序列是新冠自然變異產生的,或是多次自然重組拼合產生的,或是以自然變異+自然重組的混合方式獲得的,為什麼龐大的病毒界的無數病毒,惟獨新冠自然獲得了這一短序列,而其它病毒則無一演化出同樣的短序列?
如果新冠是自然演化、自然變異、自然重組的產物,那麼,這一病毒界中獨一無二的19核苷酸短序列,新冠是通過什麼樣的自然途徑獲得的?這一包含“RRAR”furin酶切位點的病毒界獨一無二的短序列,又是如何恰巧自然出現在擁有約30000個核苷酸位點的新冠S蛋白S1、S2亞基交界處,這一對新冠致病能力至關重要的特殊位點的?
“有據核查”最後得出如下結論:
莫德納專利基因序列和新冠病毒本身中存在短而相同的序列,並不能證明該病毒來自實驗室。這個序列可以在其他生物中找到,表明它可能是自然發生的。
“有據核查”的這一結論是否合理可信?“有據核查”是在有理“有據”地澄清、還原真相,還是在偷梁換柱、指鹿為馬,並用似是而非的表象混淆視聽、瞞天過海、欲蓋彌彰?答案不言自明。
附錄
含有新冠中19核苷酸短序列的10個細菌(或古菌)的例子:
1、Mycobacterium conspicuum(李斯特菌)JCM 14738,Genbank ID:AP022613.1;
2、Amycolatopsis(無枝菌屬) sp. YIM 10,Genbank ID:CP045480.1;
3、Persicimonas caeni strain YN101(淤泥粉紅單胞菌YN101菌株),Genbank ID:CP042468.1、CP041186.1;
4、Micromonospora(小單孢菌)sp. B006,Genbank ID:CP030865.1;
5、Micromonospora aurantiaca strain 110B 2018(酸橙小單孢菌菌株 110B 2018),Genbank ID:CP031263.1;
6、Amycolatopsis albispora strain WP1(白孢無枝菌WP1菌株),Genbank ID:CP015163.1;
7、Micromonospora(小單孢菌屬) sp. WMMA2032,Genbank ID:CP024052.1;
8、Actinopolyspora erythraea strain(嗜鹽放線多孢菌株?)YIM 90600,Genbank ID:CP022752.1;
9、Mycobacterium intracellulare subsp. yongonense strain(胞內分枝桿菌亞種 永固株)Asan 36527 ,Genbank ID:CP015965.1;
10、Micromonospora rifamycinica strain(利福黴素小單孢菌株)DSM 44983 ,Genbank ID:LT607752.1。