



| 突發武漢病毒被修改正麗致命鐵證被抓住! |
| 送交者: Pascal 2020年02月07日18:07:34 於 [五 味 齋] 發送悄悄話 |
|
武漢病毒被人為修改,石正麗致命鐵證被抓住 關於疫情來源,大體上有幾種觀點:自然界進化,實驗室泄露,敵對勢力投毒,不同群體造成網絡空前的紛爭。科普者以陰謀論駁斥病毒人工合成、敵對投毒派等觀點,而民族主義者則找尋敵對勢力,批判實驗室泄露派是被境外勢力利用,這個爭論有些亂…… 2月2日,武漢病毒所石正麗發表聲明:2019年新型冠狀病毒是大自然給人類不文明生活習慣的懲罰,我石正麗用我的生命擔保,與實驗室沒有關係。奉勸那些相信並傳播不良媒體的謠傳的人,閉上你們的臭嘴。同時轉發這個打臉消息:印度學者已經決定撤回這篇預印本文章。 2月5日,石正麗在接受財新記者採訪時再次回應爭議,希望國家專業部門來調查,以還團隊一個清白。“我自己的話沒有說服力,我不能控制別人的思想和言論。 以下為武小華回應與石正麗的爭議,特轉發。 關於我質疑石正麗研究員的朋友圈,在我睡了一個小覺後開始漫天飛,我覺得這並不是一個複雜的問題,而且相當簡單。 面對幾萬人的感染,幾萬家庭的支離破碎,幾百條人命,石研究員公然撒謊也就算了,還罵這些不幸的人活該,因為是你們自己不文明習慣的懲罰,請問這些人都是吃蝙蝠吃的嗎?荒唐!而且要質疑你的研究的科學家閉嘴,你已經喪失了最基本一個科研工作者的最基本要素:實事求是,以及一個科研工作者的社會底線:人性。 當你說出這樣的的話的時候,我真的是被你氣的咬牙切齒,那麼我就公開的把你的謊言揭露一下吧,揭露一下你的赤裸裸謊言。 1,從蝙蝠到人,新冠病毒是如何變異的?
***************************** 微信群里微民再次瘋傳留言 —— 美國病毒學家牽頭聯合中國病毒專家 就曾合作成功提取和鑑定出了一種類SARS 新型冠狀嵌合病毒,並且之後又人工製造和 培養出了一種SARS新型冠狀重組病毒。 這份研究報告由美國學者和中國學者聯合完成, 作者包括美國德克薩斯大學醫學分校微生物與 免疫學系 教授Vineet D. Menachery ,中科院 武漢病毒研究所研究員石正麗(Zhengli-Li Shi) 等人,提取發現並又製造培養了一種嵌合型 SARS樣 的新型冠狀病毒 ...... ...... 基於以上這些發現, 上述科學家們最終成功合成了一株具有感染性的 全長SHC014重組病毒, 並同時在體外和體內證實了該病毒的強大複製能力。 成功地人工合成、製造、培養出了 一種新型冠狀重組病毒!
由中國武漢開始爆發的新型冠狀病毒肺炎疫情持續引發關注,從 1 月 21 日開始證實新冠病毒肯定有“人傳人”之後,一場全民參與的抗疫戰役在中國,乃至世界正式打響。目前已知 2019 年 12 月開始,從第一起華南海鮮市場病例被證實,與SARS冠狀病毒極為類似的“不明病毒肺炎”新型冠狀病毒開始蔓延。 但是鈦媒體最近在仔細研究和整理新冠病毒相關研究時也發現,其實早在 5 年前,也就是 2015 年,美國病毒學家牽頭聯合中國病毒專家就曾合作成功提取和鑑定出了一種類SARS新型冠狀嵌合病毒,並且之後又人工製造和培養出了一種SARS新型冠狀重組病毒。這次的研究成果也被發表在了2015年的國際頂級科學雜誌《Nature》上(下稱:Nature論文)。 在今年武漢爆發的新冠病毒肺炎發生之時,回過頭來再看這份五年前的研究報告,依然頗覺觸目驚心。雖然並不能證明五年前的研究發現和本次發生的武漢肺炎病毒是完全同一的病毒,但是的確都是與Sars極為相近的新冠病毒,在今年1月,有專家認為本次“不明肺炎”疫情,傳播速度快、重病率高、難以"防控”的態勢,是世界首次發生的時候,實際上,五年前就早有發生,而病毒科學家也早有預知,並且所有預知的內容都與本次發生的疫情狀況高度相似。 這份研究報告由美國學者和中國學者聯合完成,作者包括美國德克薩斯大學醫學分校微生物與免疫學系 教授Vineet D. Menachery ,中科院武漢病毒研究所研究員石正麗(Zhengli-Li Shi)等人,提取發現並又製造培養了一種嵌合型 SARS樣 的新型冠狀病毒,根據論文所述,該病毒能讓小鼠感染上 SARS(非典肺炎),也可以證實這一病毒通過蛋白外殼與體內 RNA 進行結合,感染和傳遞到人類細胞當中,報告結論就是這種類Sars新冠狀病毒,因其高度可人際傳播的屬性,可能會給人類帶來巨大的社會風險。 五年前的預言一語成讖,同時這份研究報告也說明,本次武漢疫情可能不是偶然的,類Sars冠狀病毒的變異和傳播從未停止,但人類並未重視。正如近日,北卡羅來納州吉林斯大學全球公共衛生學院的冠狀病毒專家兼助理教授蒂莫西·謝漢(Timothy Sheahan)評價武漢新冠肺炎所說,“這不是“一次性”的病毒疫情,很可能會在未來持續發生。” 針對近期沸沸揚揚的關於武漢病毒研究所的猜測和爭議,石正麗在其朋友圈發表聲明稱:“以生命擔保,2019新冠病毒與實驗室無關,這是大自然對人類不文明生活習慣的懲罰”。
武漢病毒研究所研究員石正麗 2015年到底發生了什麼?2015年的Nature論文中闡述了一個背景,即當時在中國有一個叫做馬蹄蝠的蝙蝠種群,這是一種在岩洞棲息地里的菊頭蝠科蝙蝠群體,它們體內正在流行一種類似SARS的病毒——SHC014-CoV,而當時嚴重急性呼吸綜合徵冠狀病毒(SARS-CoV)和中東呼吸綜合徵(MERS)-CoV的出現,突出了跨物種傳播事件對人類社會的威脅,故而科學家們希望研究在馬蹄蝠中流行的這一病毒的致病潛力。 蝙蝠是許多病毒的自然宿主,包括埃博拉病毒、馬爾堡病毒,狂犬病毒、亨德拉病毒、尼帕病毒等。由於蝙蝠特殊的免疫系統,攜帶病毒卻極少出現病症。在漫長的進化歷程中,蝙蝠成為了上百種病毒的自然宿主。對於病毒溯源的研究來說,蝙蝠地位很特殊,是重點的關注對象。 中國科研人員已發現,蝙蝠體內一個被稱為“干擾素基因刺激蛋白-干擾素”的抗病毒免疫通道受到抑制,這使得蝙蝠剛好能夠抵禦疾病,卻不引發強烈的免疫反應。野生蝙蝠可能會攜帶很多病毒,但是它們都維持在一個較低的水平上。 於是,Vineet D. Menachery教授等利用SARS-CoV的反向遺傳系統,從中提取並鑑定了一種嵌合病毒,該病毒在適應小白鼠的SARS-CoV主幹中可表達出蝙蝠冠狀病毒SHC014的刺突。結果表明,該新型冠狀病毒能利用SARS的人類細胞受體——血管緊張素轉換酶II(ACE2)的多個同源基因,在人類呼吸道原代細胞中有效複製,並在體外獲得與SARS傳染性同等的效果。 此外,體內實驗證明,該嵌合病毒被複製在小白鼠肺部後,可導致明顯的發病徵狀。評估顯示,現有的基於SARS的免疫治療和預防模式的效果不佳;單克隆抗體和疫苗方法均未能利用這種新的刺突蛋白來中和免疫CoVs感染。 基於以上這些發現,上述科學家們最終成功合成了一株具有感染性的全長SHC014重組病毒,並同時在體外和體內證實了該病毒的強大複製能力。研究結果表明,這種病毒完全可重現SARS-CoV(SARS冠狀病毒)的傳播風險。 Nature論文三種範例所證明的新冠病毒傳播原理,與武漢疫情高度類似以下都來自鈦媒體編輯翻譯並整理了上述2015年《Nature》論文中的主要分析內容,傳播原理的確與本次武漢新冠肺炎類似,很多結果也類似,例如關於致病性不會比Sars冠狀病毒增強、人傳人潛在風險極大(傳染性大)、老年動物(實驗主要以動物進行實驗)更易感等。 高致病性冠狀病毒SARS冠狀病毒的出現,預示着嚴重呼吸疾病跨物種傳播的新時代。全球化為病毒的快速傳播帶來基礎,對全球經濟有着巨大影響。 在SARS席捲全球後,甲型流感亞型病毒H5N1、H1N1和H7N9以及MERS冠狀病毒(中東呼吸綜合徵)開始出現,並且可以同樣通過動物傳染人類,對當地人口帶來了死亡的陰影以及經濟損失。 儘管公共衛生措施控制了SARS冠狀病毒的爆發,但最近的宏基因組研究通過識別近期在中國馬蹄蝠中大面積流行的類SARS冠狀病毒的病毒序列,發現這些由蝙蝠攜帶的病毒有可能在未來帶來更多的威脅。 但是,病毒序列帶來的數據只能為將來識別以及預防類似流行性病毒提供非常有限的洞見。因此,為了分析蝙蝠攜帶的冠狀病毒引起流行性傳播的可能性(也即是傳染人類的可能性),我們提取了中國馬蹄蝠攜帶的RsSHC014冠狀病毒序列中可造成人畜傳染的冠狀病毒刺穿蛋白,使用適應小白鼠的SARS冠狀病毒主鏈培育出了一種嵌合病毒。 培育出來的混合病毒讓我們能夠評估該刺穿蛋白在沒有經過自然適應性變異的情況下感染人類的能力。通過這個方法,我們對人類氣道細胞在活體狀態下由SHC014冠狀病毒導致的感染特徵進行了分析,測試了現有的免疫和治療方法對SHC014冠狀病毒的作用。這個研究方法能對宏基因組學數據進行解讀,幫助預防未來可能會出現的病毒,並未將來疫情爆發的可能性做好準備。 SHC014和相關的RsWIV1冠狀病毒的序列顯示,該類冠狀病毒為SARS冠狀病毒(圖示1a,b)的近親。但是,SHC014和SARS冠狀病毒在結合人類的14殘基和SARS冠狀病毒受體中存在差異。受體包括5個決定宿主範圍的殘基:Y442、L472、N479、T487和Y491 。在WIV1中,其中三個殘基與Urbani SARS冠狀病毒不同,按原本預期,它們不會改變和ACE2的結合。 這一情況由兩個假型化實驗得到證實。其中一個實驗測試了慢病毒屬WIV1刺穿蛋白進入細胞釋放血管緊張素轉換酶II(AC2)的能力。另一實驗為複製WIV1冠狀病毒(ref. 1)的試管實驗。在SHC014中存在14個與ACE2有相互作用的殘基,其中有7個殘基和SARS冠狀病毒的不一樣,這包括了所有5個對決定宿主範圍起關鍵作用的殘基。 這些變化,以及釋放SHC014刺穿蛋白進入細胞的慢病毒假型實驗,表明SHC014刺穿蛋白無法與人類的ACE2進行結合。 但是,有研究發現,其他相關的SARS冠狀病毒株存在類似變化,這意味着我們需要做進一步的功能測試才能做出更好的判斷。因此,我們基於複製功能強、實應小白鼠的SARS冠狀病毒主鏈合成了SHC014刺穿蛋白(該嵌合冠狀病毒在後文統稱為SHC014-MA15),以研究小白鼠的發病機理和疫苗研究。與現有分子建模和假型實驗得出的預測不同,SHC014-MA15保持了活性,並且在Vero細胞中複製致高滴度。 類似於SARS病毒,SHC014-MA15同樣需要功能正常的ACE2分子才能進入細胞,並且可以使用與人類、果子狸和蝙蝠直系同源ACE2。為了測試SHC014刺穿蛋白感染人類氣道的能力,我們研究了人類上皮氣道細胞系Calu-3 2B4對感染的敏感性,發現SHC014-MA15具有強大複製能力,與Urbani SARS病毒相當。 除此之外,人類主要氣道上皮(HAR)培養物被感染,兩種病毒都展現出了強大的複製能力。總之,這些數據證實了帶有SHC014的病毒感染人類氣道細胞的能力,並顯示出了SHC014冠狀病毒具備跨物種傳播的潛在威脅。
圖|SARS 樣的冠狀病毒在人類氣道細胞中複製並產生體內的發病機理 為了評估SHC014次突蛋白在體內介導感染中的作用,我們用104個SARS-MA15或SHC014-MA15空斑形成單位 (p.f.u.)感染了10周大小的BALB/c小白鼠(圖1e-h)。 感染SARS-MA15的白鼠在感染後第4天出現體重迅速下降和死亡現象(d.p.i.);而感染了SHC014-MA15的小白鼠體重下降顯著(10%),但無死亡(圖1e)。 對病毒複製的檢測顯示,感染了SARS-MA15或SHC014-MA15的小鼠肺部病毒滴度幾乎相同(圖1f)。感染了SARS-MA15的小白鼠的肺的末梢細支氣管和肺實質(圖1g)均有較強程度的染色(圖1g),而被SHC014-MA15感染的小白鼠的肺抗原染色較弱(圖1h);與此相反,在軟組織或整體組織評分中未發現抗原染色缺陷,這表示肺部組織對SHC014-MA15的感染情況有異(補充表2)。 感染SARS-MA15的動物體重迅速下降,並死於感染(補充圖3a,b)。感染SHC014-MA15可導致動物體重顯著下降,但致死率極低。在年輕小白鼠中觀測到的組織學和抗原染色模式,在年長些的白鼠中也可觀測到(補充表3)。我們使用Ace2−−老鼠進行實驗,排除了SHC014-MA15通過另一種感染受體傳播的可能性。 該種情況下,小白鼠沒有體重下降,或被SHC014-MA15感染後的抗原染色現象(補充圖4 a, b和補充表2)。以上這些數據表明,具有SHC014刺突的病毒能夠在CoV病毒主幹的條件下致使小白鼠體重減輕。 考慮到埃博拉單克隆抗體療法(例如 ZMApp 10)的臨床功效,我們接下來試圖確定 SARS-CoV 單克隆抗體對抗 SHC014-MA15 感染的功效。廣泛中和針對 SARS 冠狀病毒刺突蛋白已經被先前報道,並且是免疫試劑可能人類單克隆抗體。 我們檢查了這些抗體對病毒複製的影響(表示為病毒複製的抑制百分比),發現野生型 SARS-CoV Urbani 在相對較低的抗體濃度下被所有四種抗體強烈中和,SHC014-MA15的中和效果有所不同。通過噬菌體展示產生的和逃避突變型的抗體,僅實現抑制 SHC014-MA15 複製(如圖 2a)。同樣,源自 SARS-CoV 感染患者的記憶 B 細胞的抗體 230.15 和 227.14 也未能阻止SHC014-MA15複製(圖 2b,c)。 對於所有三種抗體, SARS 和 SHC014 尖峰氨基酸序列之間的差異對應於 SARS-CoV 逃逸突變體(fm6 N479R; 230.15 L443V; 227.14 K390Q / E)中發現的直接或相鄰殘基變化,這可能解釋了抗體的缺失針對SHC014的中和活性。 最後,單克隆抗體 109.8 能夠實現 SHC014-MA15 的 50% 中和,但僅在高濃度(10μg/ ml)時(圖2d)。總之,結果表明,針對 SARS-CoV 的廣泛中和抗體可能僅對新興 SARS 樣 CoV 菌株(如 SHC014)具有邊際功效。
圖|SARS-CoV單克隆抗體對SARS樣CoV的療效折線圖 為了評估現有疫苗抵抗 SHC014-MA15 感染的功效,研究人員用雙重滅活的完整 SARS-CoV(DIV)疫苗接種了老年小鼠。先前的工作表明,這種疫苗接種方式可以中和並保護年輕小鼠免受同源病毒的攻擊;然而,該疫苗未能保護其中還觀察到增強的免疫病理的老年動物,這表明由於疫苗接種,動物受到了傷害。在這裡,我們發現 DIV 不能在體重減輕或病毒滴度方面提供 SHC014-MA15 的抗攻擊保護(圖5a,b)。與其他異源組 CoV 的先前報告一致。此外,來自接種 DIV 的老年小鼠的血清也未能中和 SHC014-MA15(如圖5c)。 值得注意的是, DIV 疫苗接種導致了強大的免疫病理和嗜酸性粒細胞增多。這些結果證實,DIV 疫苗不能預防 SHC014 感染,並且可能增加老年接種組的疾病。 與用 DIV 疫苗接種相比,將 SHC014-MA15 用作減毒活疫苗顯示了針對 SARS-CoV 攻擊的潛在交叉保護作用。論文當中的研究人員表示,其用 10 4pfu 的 SHC014-MA15 感染幼鼠,並觀察了 28 天。然後,在第 29 天,他們用 SARS-MA15 攻擊了小鼠。 儘管在 SHC014-MA15 感染後28 天內產生的抗血清,只有極少的 SARS-CoV 中和反應,但先前用高劑量 SHC014-MA15 感染的小鼠可防禦致命劑量的 SARS-MA15 攻擊。 在沒有二級抗原升壓的情況下,28 dpi 表示的抗體滴度的預期峰值,意味着有將隨時間而減少針對 SARS-CoV 的保護。在體重減輕和病毒複製方面,在衰老的 BALb/c 小鼠中觀察到了類似的結果,表明用致死劑量的SARS-CoV攻擊具有保護作用。然而,在某些老年動物中,SHC014-MA1 5感染劑量為 10 4pfu,導致體重減輕和致死率> 10%。 研究人員發現,以較低劑量的 SHC014-MA15(100 pfu)進行疫苗接種不會誘導體重減輕,但也無法保護老年動物免受 SARS-MA15 致命劑量的攻擊。總之言之,根據論文的說法,數據表明,SHC014-MA15 攻擊可能通過保守的表位賦予針對 SARS-CoV 的交叉保護作用,但所需劑量可誘發發病機理,並不能用作減毒疫苗。 確定 SHC014 尖峰具有介導人類細胞感染並引起小鼠疾病的能力後,我們接下來基於用於SARS-CoV的方法合成了全長SHC014-CoV感染性克隆。在Vero細胞中複製顯示SHC014-CoV相對於SARS-CoV沒有缺陷;然而,在感染後24小時和48小時,原代HAE培養物中SHC014-CoV顯着減弱(P<0.01)。 與SARS-CoV Urbani相比,小鼠的體內感染沒有顯示出明顯的體重減輕,但在全長SHC014-CoV感染的肺部顯示出病毒複製減少。總之,這些結果確定了全長SHC014-CoV的生存力,但表明其複製必須與人類呼吸道細胞和小鼠中流行的SARS-CoV的複製相一致,需要進一步的適應。
圖|SHC014-CoV在人呼吸道中複製,但缺乏流行性SARS-CoV的毒力 在 SARS 冠狀病毒流行期間,人們很快發現了棕櫚科動物和人類冠狀病毒之間的聯繫。基於這一發現,有兩種不同觀點範例表述。其中,第一種觀點認為,流行性 SARS-CoV 起源於蝙蝠病毒,躍遷至小窩並在受體結合域(RBD)內引入了變化,以改善與麝貓 Ace2 的結合,隨後在活畜市場上接觸人,使人感染了麝香毒株,而該麝香毒株又適合成為流行毒株。 但是,根據發育周期以及數據分析表明,早期人類SARS菌株與蝙蝠菌株的關係似乎比靈貓菌株更為緊密。因此,第二種觀點認為蝙蝠直接傳播導致SARS-CoV的出現,而棕櫚科動物則是繼發宿主和持續感染的宿主。對於這兩種範例,都認為必須在二級宿主中適應峰值,因為大多數突變預計會在RBD內發生,從而有助於改善感染。兩種理論都暗示蝙蝠冠狀病毒的庫是有限的,宿主範圍的突變既是隨機的又是罕見的,從而降低了人類未來出現突發事件的可能性。
圖|冠狀病毒的出現(傳播)範例 儘管Nature論文表示,這一新的研究,並未打破上述說法與範例,但也的確提出了第三種範式,這一論文的作者團隊認為,蝙蝠 CoV 病毒細胞為了保持“平衡”的刺突蛋白,能夠在不突變的情況下感染人類。通過在SARS-CoV主鏈中包含SHC014刺突的嵌合病毒在人氣道培養物中和在沒有RBD適應的小鼠中引起強烈感染的能力可以說明這一假設。 加上以前鑑定的致病冠狀病毒主鏈的觀察,上述論文研究結果表明,SARS 樣菌株所急需的原材料目前正在動物水庫中流通。值得注意的是,儘管全長SHC014-CoV可能需要額外的骨架適應性來介導人類疾病,但有記錄的CoV家族中的高頻重組事件強調了未來出現的可能性和進一步準備的必要性。 迄今為止,動物種群的基因組學篩選已主要用於鑑定暴發環境中的新型病毒。這裡的方法將這些數據集擴展為檢查病毒出現和治療功效的問題。我們認為,具有SHC014尖峰的病毒由於其在原代人類氣道培養物中複製的能力而成為潛在的威脅,這是人類疾病的最佳可用模型。另外,在小鼠中觀察到的發病機制表明含有SHC014的病毒能夠在沒有RBD適應的情況下在哺乳動物模型中引起疾病。 值得注意的是,相對於SARS-CoV Urbani,與SARS-MA15相比,肺中的向異性和HAE培養物中全長SHC014-CoV的衰減相對於SARS-CoV Urbani而言,提示了ACE2結合以外的因素-包括穗突性,受體生物利用度或拮抗宿主免疫反應中的一部分可能有助於出現。然而,需要對非人類靈長類動物進行進一步測試,以將這些發現轉化為人類致病潛能。重要的是,現有治療方法的失敗定義了進一步研究和開發治療方法的關鍵需求。有了這些知識,就可以產生監視程序,診斷試劑和有效的治療方法,從而防止出現組別特異性CoV,例如SHC014,並且可以將其應用於維護相似異質庫的其他CoV分支。 在以前出現的模型的基礎上,研究人員認為,新型冠狀病毒並沒有增強致病性。作者表示,預計不會產生嵌合病毒(例如SHC014-MA15)會增加致病性的情況。雖然SHC014-MA15相對於其親本小鼠適應的SARS-CoV減毒,但類似的研究檢查了MA15骨架內野生型Urbani尖峰的CoV的致病性,顯示小鼠無體重減輕,病毒複製減少。因此,相對於Urbani峰值–MA15 CoV,SHC014-MA15的發病機理有所改善。 基於這些發現,科學評論小組可能認為類似的研究基於無法冒險進行的循環株構建嵌合病毒,因為不能排除哺乳動物模型中致病性的增加。再加上對小鼠適應株的限制以及使用逃逸突變體開發單克隆抗體的研究,對CoV出現和治療功效的研究可能會嚴重受限。這些數據和限制加在一起,代表了新冠病毒研究關注的十字路口。在制定前進的政策時,必須權衡準備和緩解未來爆發的潛力與創造更多危險病原體的風險。 總體而言,研究人員利用實驗和分析,對已使用宏基因組學數據來識別由循環蝙蝠SARS狀CoV SHC014構成的潛在威脅。由於嵌合的SHC014病毒在人氣道培養物中複製的能力,在體內引起發病機理並逃避當前的治療方法,因此需要針對循環的SARS樣病毒的監視和改進的治療方法。我們的方法還可以利用宏基因組學數據來預測病毒的出現,並將這些知識應用於準備治療未來出現的病毒感染。 培養並製造出新冠病毒的實驗過程以下是鈦媒體編輯整理了上述論文中科學家們培養並製造出一種新型冠狀病毒的實驗過程,這一過程被稱為“病毒,細胞,體外感染和噬菌斑測定方案”。 科學家將從美國陸軍傳染病研究所獲得的野生型 SARS-CoV(Urbani),適應小鼠的 SARS-CoV(MA15)和嵌合型 SARS 型 CoV 病毒細胞在 Vero E6 培養基細胞上。據悉,這一細胞是根據美國病毒學家杜爾貝科(Dulbecco,Renato) 所研製出改良的 Eagle's 培養基上生長。 另外,獲得到的病毒細胞組織,是通過之前研究的信息進行的,其中包括表達 ACE2 直向同源物的 DBT 細胞(Baric實驗室,來源未知),這些都在先前被描述為人類和麝貓感染的細胞組織,ACE2 序列是基於從菊頭蝠,就是蝙蝠 DBT 細胞當中提取,並從中建立新的病毒培養。 作者表示,這一實驗本身是偽型實驗過程,與使用基於 HIV 的偽病毒實驗類似,先前使用武漢病毒研究所提供的 ACE2 直系表達的同源物種的 HeLa 細胞進行檢查,HeLa 細胞在最低必需培養基補充有 10% FCS(Gibco公司,CA)以及 MEM(Gibco公司,CA)中生長,得出下圖在Vero E6,DBT,將Calu-3和2B4原代人呼吸道上皮細胞的生長曲線。 實驗物,也就是仿製的肺,則是在北卡羅來納大學機構審查委員會批准下採購的,而這種肺並不是活體的肺,是在 HAE 培養的人肺。該 HAE 培養物代表高度分化的人氣道上皮,其中包含纖毛和非纖毛上皮細胞以及杯狀細胞,培養物應在氣液界面上生長數周,才拿到該實驗當中。 而後,將細胞用 PBS 溶液進行洗滌,並用病毒接種或在 37℃ 的 PBS 溶液中模擬稀釋 40 分鐘。將細胞洗滌 3 次,並加入新鮮培養基以表示時間 “0”。在每個所述的時間點收穫三個或更多的生物重複樣品,所有病毒的培養均在生物安全級別(BSL)為 3 級的實驗室中進行。 人員和環境方面,將冗餘的生物細胞放進對應的安全櫃中,並有電扇進行散熱。所有人員都穿着特別的衛生強化防護服,為了實驗過程的健康,相關科研人員使用 3M 電動呼吸器(Breathe Easy,3M)進行呼吸,並嚴格佩戴雙手套,使得實驗在安全狀況下進行。 序列聚類和結構建模 實驗中,從 Genbank 或 Pathosystems 資源整合中心(PATRIC)下載具有代表性的 CoV S1 結構域的全長基因組序列和氨基酸序列,並與 ClustalX 進行比對,通過系統分析使用 100 個自舉法或使用 PhyML(https://code.google.com/p/phyml/)封裝。 事實上,這一過程主要是將數據進行比對,對病毒的未來變異進行管控,形成一定的數據模型。 接着,科研人員使用 PhyML 軟件包生成最大可能性數值樹。比例尺代表核苷酸取代。僅標記引導程序支持高於 70% 的節點。該樹顯示 CoV 分為三個不同的系統發育組,分別定義為 α-CoV,β-CoV 和 γ-CoV。對 於β-CoV,經典子群群集標記為 2a,2b,2c 和 2d,對於 α-CoV,標記為 1a 和 1b。使用 Modeller(Max Planck Institute Bioinformatics Toolkit)生成結構模型,以基於晶體結構 2AJF(蛋白質數據庫)生成 SARS RBD 的 SHC014 和 Rs3367 與 ACE2 的同源性模型。在 MacPyMol(1.3版)中可視化和操作同源模型。 SARS樣嵌合病毒的構建 如前所述,野生型和嵌合病毒均來自 SARS-CoV Urbani 或相應的小鼠適應性(SARS-CoV MA15)感染性克隆(ic)病毒細胞。通過提取含有 SHC014 刺突序列的質粒,並連接到 MA15 感染性克隆的 E 和 F 質粒中。設計該克隆並從 Bio Basic 購買六種連續 cDNA,並使用側接獨特的 II 類限制性核酸內切酶位點(BglI)的公開序列。 之後,研究人員對其擴增,切除,連接和純化含有野生型,嵌合 SARS-CoV 和 SHC014-CoV 基因組片段的質粒。然後進行體外轉錄反應以合成全長基因組 RNA,如先前所述將其轉染到 Vero E6 細胞中。收穫轉染細胞的培養基,並用作後續實驗的種子庫。在用於這些研究之前,通過序列分析證實了嵌合和全長病毒。嵌合突變體和全長 SHC014-CoV 的合成構建,這一新的 SARS樣嵌合病毒標準,需要得到北卡羅萊納大學機構生物安全委員會和關注雙重用途研究委員會的批准。 小鼠體內感染 事實上,這份實驗需要根據人體,來了解抵抗力以及病毒形成活動,多數據下,形成小鼠體內感染的數據分析。 論文中稱,其從 Harlan Laboratories 訂購了雌性、10 周齡和 12 個月大的 BALb/c 白變種實驗室老鼠。將動物帶入 BSL3 實驗室,並使其在感染前適應 1 周。對於感染和減毒活疫苗接種,將用氯胺酮和甲苯噻嗪的混合物對小鼠進行麻醉,並在鼻腔感染時用 50μl 磷酸鹽緩衝液(PBS)或稀釋的病毒經鼻內感染,每個時間點,每隻三或四隻小鼠,感染組每劑量都按照條件定量。對於個別小鼠,研究人員可以酌情決定是否將小鼠數據排除在外,包括不吸入全部劑量,鼻腔冒泡或通過口部感染引起的感染。 感染後,在任何動物實驗中均未使用致盲法,且動物未隨機分組。對小鼠進行接種疫苗,通過足墊注射為年輕和老年小鼠接種 20μl 的含明礬或模擬 PBS 的 0.2μg 雙滅活 SARS-CoV 疫苗;然後在 22 天后以相同的方案加強小鼠免疫體質。對於所有組,按照實驗方案,在實驗過程中每天監測動物的疾病臨床體徵(駝背,毛皮鬆動和活動減少)。在頭 7 天裡,每天監測體重是否減輕,此後繼續進行體重監測,直到動物恢復至初始初始體重或連續 3 天顯示體重增加為止。所有體重下降超過其初始體重 20% 的小鼠都被餵食並每天監測多次,只要它們處於 20% 的臨界值以下即可。 按照實驗方案,立即處死體重超過其初始體重 30% 的小鼠。根據研究人員的說法,任何被認為垂死,或不太可能恢復的小鼠,將使用異氟烷過量進行安樂死,並通過頸脫位確認死亡,這種處理方式使用 UNC 機構動物護理和使用委員會(IACUC)批準的方案。 組織學分析 在實驗結束後,取出左肺,浸沒在 10% 福爾馬林緩衝液(Fisher)中,不充氣 1 周。將組織包埋在石蠟中,並由 UNC Lineberger 綜合癌症中心組織病理學核心設施製備 5μm 切片。為了確定抗原染色的程度,使用市售的多克隆 SARS-CoV 抗核衣殼抗體(Imgenex)對切片進行病毒抗原染色,並以盲法方式對氣道和實質進行染色,使用帶有 Olympus DP71 相機的 Olympus BX41 顯微鏡捕獲圖像。 病毒中和測定和統計分析 在這一部分當中,噬斑減少中和效價測定法與先前表徵針對 SARS-CoV 的抗體進行。簡而言之,就是中和抗體或血清,並將其進行兩次連續稀釋,與 100pfu 不同感染性克隆 SARS-CoV 菌株在 37°C 溫度下孵育 1 小時。然後將病毒和抗體以 5×10 5 孔 Vero E6 細胞添加到另一孔板中,並重複多次(n≥2)。 在 37°C 下孵育 1 小時後,研究人員需要在培養基中鋪上 3 ml 0.8% 瓊脂糖。在將板在 37℃ 下孵育 2 天,用中性紅染色 3 小時並計數噬菌斑。計算噬菌斑減少的百分比為(1-((帶有抗體的噬菌斑數量/沒有抗體的噬菌斑數量))×100。 實驗結束後進行統計分析,所有實驗均與兩個實驗組(兩種病毒,或已接種和未接種的隊列)進行對比。因此,病毒的顯著差異是通過在各個時間點進行檢驗確定的。最終的數據以正態分布,在每個比較的組別中,放置出相似的方差。(本文首發鈦媒體App) 原論文地址: https://www.nature.com/articles/nm.3985#auth-14 參考來源: https://microbiology.utmb.edu/faculty/vineet-d-menachery-phd https://www.the-scientist.com/news-opinion/lab-made-coronavirus-triggers-debate-34502 https://www.nature.com/articles/d41586-020-00262-7
實驗室里人工合成製造培養 的冠狀病毒引發巨大爭議
Ralph Baric, an infectious-disease researcher at the University of North Carolina at Chapel Hill, last week (November 9) published a study on his team’s efforts to engineer a virus with the surface protein of the SHC014 coronavirus, found in horseshoe bats in China, and the backbone of one that causes human-like severe acute respiratory syndrome (SARS) in mice. The hybrid virus could infect human airway cells and caused disease in mice, according to the team’s results, which were published in Nature Medicine. The results demonstrate the ability of the SHC014 surface protein to bind and infect human cells, validating concerns that this virus—or other coronaviruses found in bat species—may be capable of making the leap to people without first evolving in an intermediate host, Nature reported. They also reignite a debate about whether that information justifies the risk of such work, known as gain-of-function research. “If the [new] virus escaped, nobody could predict the trajectory,” Simon Wain-Hobson, a virologist at the Pasteur Institute in Paris, told Nature. In October 2013, the US government put a stop to all federal funding for gain-of-function studies, with particular concern rising about influenza, SARS, and Middle East respiratory syndrome (MERS). “NIH [National Institutes of Health] has funded such studies because they help define the fundamental nature of human-pathogen interactions, enable the assessment of the pandemic potential of emerging infectious agents, and inform public health and preparedness efforts,” NIH Director Francis Collins said in a statement at the time. “These studies, however, also entail biosafety and biosecurity risks, which need to be understood better.” Baric’s study on the SHC014-chimeric coronavirus began before the moratorium was announced, and the NIH allowed it to proceed during a review process, which eventually led to the conclusion that the work did not fall under the new restrictions, Baric told Nature. But some researchers, like Wain-Hobson, disagree with that decision. The debate comes down to how informative the results are. “The only impact of this work is the creation, in a lab, of a new, non-natural risk,” Richard Ebright, a molecular biologist and biodefence expert at Rutgers University, told Nature. But Baric and others argued the study’s importance. “[The results] move this virus from a candidate emerging pathogen to a clear and present danger,” Peter Daszak, president of the EcoHealth Alliance, which samples viruses from animals and people in emerging-diseases hotspots across the globe, told Nature. *******************************
bats are an important source of highly lethal zoonotic viruses, such as Hendra, Nipah, Ebola and Marburg viruses16. Here we report on a series of fatal swine disease outbreaks in Guangdong province, China, approximately 100 km from the location of the purported index case of SARS. Most strikingly, we found that the causative agent of this swine acute diarrhoea syndrome (SADS) is a novel HKU2-related coronavirus that is 98.48% identical in genome sequence to a bat coronavirus, which we detected in 2016 in bats in a cave in the vicinity of the index pig farm. This new virus (SADS-CoV) originated from the same genus of horseshoe bats (Rhinolophus) as SARS-CoV. From 28 October 2016 onwards, a fatal swine disease outbreak was observed in a pig farm in Qingyuan, Guangdong province, China, very close to the location of the first known index case of SARS in 2002, who lived in Foshan (Extended Data Fig. 1a). Porcine epidemic diarrhoea virus (PEDV, a coronavirus) had caused prior outbreaks at this farm, and was detected in the intestines of deceased piglets at the start of the outbreak. However, PEDV could no longer be detected in deceased piglets after 12 January 2017, despite accelerating mortality (Fig. 1a), and extensive testing for other common swine viruses yielded no results (Extended Data Table 1). These findings suggested that this was an outbreak of a novel disease. Clinical signs are similar to those caused by other known swine enteric coronaviruses17, 18 and include severe and acute diarrhoea and acute vomiting, leading to death due to rapid weight loss in newborn piglets that are less than five days of age. Infected piglets died 2–6 days after disease onset, whereas infected sows suffered only mild diarrhoea and most sows recovered within two days. The disease caused no signs of febrile illness in piglets or sows. The mortality rate was as high as 90% in piglets that were five days or younger, whereas in piglets that were older than eight days, the mortality dropped to 5%. Subsequently, SADS-related outbreaks were found in three additional pig farms within 20–150 km of the index farm (Extended Data Fig. 1a) and, by 2 May 2017, the disease had caused the death of 24,693 piglets at these four farms (Fig. 1a). In farm A alone, 64% (4,659 out of 7,268) of all piglets that were born in February died. The outbreak has abated, and measures that were taken to control SADS included separation of sick sows and piglets from the rest of the herd. A qPCR test described below was used as the main diagnostic tool to confirm SADS-CoV infection. Fig. 1: Detection of SADS-CoV infection in pigs in Guangdong, China. a, Records of daily death toll on the four farms from 28 October 2016 to 2 May 2017. b, Detection of SADS-CoV by qPCR. The y axis shows the log(copy number per 106 copies of 18S rRNA). n = 12 sick piglets, 5 sick sows, 16 recovered sows and 10 healthy piglets. c, Tissue distribution of SADS-CoV in diseased pigs. n = 3. Data are mean ± s.d.; dots represent individual values. d, Detection of SADS-CoV antibodies. n = 46 sows from whom serum was first taken in the first three weeks of the outbreak (First bleed), n = 8 sows from whom serum was taken again (Second bleed) at more than one month after the onset of the outbreak, n = 8 sera from healthy pig controls, n = 35 human sera from pig farmers. A sample collected from the small intestine of a diseased piglet was analysed by metagenomics analysis using next-generation sequencing (NGS) to identify potential aetiological agents. Of the 15,256,565 total reads obtained, 4,225 matched sequences of the bat CoV HKU2, which was first detected in Chinese horseshoe bats in Hong Kong and Guangdong province, China19. By de novo assembly and targeted PCR, we obtained a 27,173-bp CoV genome that shared 95% sequence identity to HKU2-CoV (GenBank accession number NC_009988). Thirty-three full genome sequences of SADS-CoV were subsequently obtained (8 from farm A, 5 from farm B, 11 from farm C and 9 from farm D) that were 99.9% identical to each other (Supplementary Table 1). Using qPCR targeting the nucleocapsid gene (Supplementary Table 2), we detected SADS-CoV in acutely sick piglets and sows, but not in recovered or healthy pigs on the four farms, nor in nearby farms that showed no evidence of SADS. The virus replicated to higher titres in piglets than in sows (Fig. 1b). SADS-CoV displayed tissue tropism of the small intestine (Fig. 1c), as observed for other swine enteric coronaviruses20. Retrospective PCR analysis revealed that SADS-CoV was present on farm A during the PEDV epidemic, where the first strongly positive SADS-CoV sample was detected on 6 December 2016. From mid-January onwards, SADS-CoV was the dominant viral agent detected in diseased animals (Extended Data Fig. 1b). It is possible that the presence of PEDV early in the SADS-CoV outbreak may have somehow facilitated or enhanced spillover and amplification of SADS. However the fact that the vast majority of piglet mortality occurred after PEDV infection had become undetectable suggests that SADS-CoV itself causes a lethal infection in pigs that was responsible for these large-scale outbreaks, and that PEDV does not directly contribute to its severity in individual pigs. This was supported by the absence of PEDV and other known swine diarrhoea viruses during the peak and later phases of the SADS outbreaks in the four farms (Extended Data Table 1). We rapidly developed an antibody assay based on the S1 domain of the spike (S) protein using a luciferase immunoprecipitation system21. Because SADS occurs acutely and has a rapid onset in piglets, serological investigation was conducted only in sows. Among 46 recovered sows tested, 12 were seropositive for SADS-CoV within three weeks of infection (Fig. 1d). To investigate possible zoonotic transmission, serum samples from 35 farm workers who had close contact with sick pigs were also analysed using the same luciferase immunoprecipitation system approach and none were positive for SADS-CoV. Although the overall genome identity of SADS-CoV and HKU2-CoV is 95%, the S gene sequence identity is only 86%, suggesting that the previously reported HKU2-CoV is not the direct progenitor of SADS-CoV, but that they may have originated from a common ancestor. To test this hypothesis, we developed a SADS-CoV-specific qPCR assay based on its RNA-dependent RNA polymerase (RdRp) gene (Supplementary Table 2) and screened 591 bat anal swabs collected between 2013 and 2016 from seven different locations in Guangdong province (Extended Data Fig. 1a). A total of 58 samples (9.8%) tested positive (Extended Data Table 2), all were from Rhinolophus spp. bats that are also the natural reservoir hosts of SARS-related coronaviruses3,4,5,6,7,8,9,10. Four complete genome sequences with the highest RdRp PCR-fragment sequence identity to that of SADS-CoV were determined by NGS. They are very similar in size (27.2 kb) compared to SADS-CoV (Fig. 2a) and we tentatively call them SADS-related coronaviruses (SADSr-CoV). Overall sequence identity between SADSr-CoV and SADS-CoV ranges from 96 to 98%. Most importantly, the S protein of SADS-CoV shared more than 98% sequence identity with sequences of two of the SADSr-CoVs (samples 162149 and 141388), compared to 86% with HKU2-CoV. The major sequence differences among the four SADSr-CoV genomes were found in the predicted coding regions of the S and NS7a and NS7b genes (Fig. 2a). In addition, the coding region of the S protein N-terminal (S1) domain was determined from 19 bat SADSr-CoVs to enable more detailed phylogenetic analysis. Fig. 2: Genome and phylogenetic analysis of SADS-CoV and SADSr-CoV. a, Genome organization and comparison. Colour-coding for different genomic regions as follows. Green, non-structural polyproteins ORF1a and ORF1b; yellow, structural proteins S, E, M and N; blue, accessory proteins NS3a, NS7a and NS7b; Orange, untranslated regions. The level of sequence identity of SADSr-CoV to SADS-CoV is illustrated by different patterns of boxes: Solid colour, highly similar; Dotted fill, moderately similar; Dashed fill, least similar. b, Phylogenetic analysis of 57 S1 sequences (33 from SADS-CoV and 24 from SADSr-CoV). Different colours represent different host species as shown on the left. Scale bar, nucleotide substitutions per site. The phylogeny of S1 and the full-length genome revealed a high genetic diversity of alphacoronaviruses among bats and strong coevolutionary relationships with their hosts (Fig. 2b and Extended Data Fig. 2), and showed that SADS-CoVs were more closely related to SADSr-CoVs from Rhinolophus affinis than from Rhinolophus sinicus, in which HKU2-CoV was found. Both phylogenetic and haplotype network analyses demonstrated that the viruses from the four farms probably originated from their reservoir hosts independently (Extended Data Fig. 3), and that a few viruses might have undergone further genetic recombination (Extended Data Fig. 4). However, molecular clock analysis of the 33 SADS-CoV genome sequences failed to establish a positive association between sequence divergence and sampling date. Therefore, we speculate that either the virus was introduced into pigs from bats multiple times, or that the virus was introduced into pigs once, but subsequent genetic recombination disturbed the molecular clock. For viral isolation, we tried to culture the virus in a variety of cell lines (see Methods for details) using intestinal tissue homogenates as starting material. Cytopathogenic effects were observed in Vero cells only after five passages (Extended Data Fig. 5a, b). The identity of SADS-CoV was verified in Vero cells by immunofluorescence microscopy (Extended Data Fig. 5c, d) and by whole-genome sequencing (GenBank accession number MG557844). Similar results were obtained by other groups22, 23. Known coronavirus host cell receptors include angiotensin-converting enzyme 2 (ACE2) for SARS-related CoV, aminopeptidase N (APN) for certain alphacoronaviruses, such as human (H)CoV-229E, and dipeptidyl peptidase 4 (DPP4) for Middle East respiratory syndrome (MERS)-CoV24,25,26. To investigate the receptor usage of SADS-CoV, we tested live or pseudotyped SADS-CoV infection on HeLa cells that expressed each of the three molecules. Whereas the positive control worked for SARS-related CoV and MERS-CoV pseudoviruses, we found no evidence of enhanced infection or entry for SADS-CoV, suggesting that none of these receptors functions as a receptor for virus entry for SADS-CoV (Extended Data Table 3). To fulfill Koch’s postulates for SADS-CoV, two different types of animal challenge experiments were conducted (see Methods for details). The first challenge experiment was conducted with specific pathogen-free piglets that were infected with a tissue homogenate of SADS-CoV-positive intestines. Two days after infection, 3 out of 7 animals died in the challenge group whereas 4 out of 5 survived in the control group. Incidentally, the one piglet that died in the control group was the only individual that did not receive colostrum due to a shortage in the supply. It is thus highly likely that lack of nursing and inability to access colostrum was responsible for the death (Extended Data Table 4). For the second challenge, healthy piglets were acquired from a farm in Guangdong that had been free of diarrheal disease for a number of weeks before the experiment, and were infected with the cultured isolate of SADS-CoV or tissue-culture medium as control. Of those inoculated with SADS-CoV, 50% (3 out of 6) died between 2 and 4 days after infection, whereas all control animals survived (Extended Data Table 5). All animals in the infected group suffered watery diarrhoea, rapid weight loss and intestinal lesions (determined after euthanasia upon experiment termination, Extended Data Tables 4, 5). Histopathological examination revealed marked villus atrophy in SADS-CoV inoculated farm piglets four days after inoculation but not in control piglets (Fig. 3a, b) and viral N protein-specific staining was observed mainly in small intestine epithelial cells of the inoculated piglets (Fig. 3c, d). Fig. 3: Immunohistopathology of SADS-CoV infected tissues. a–d, Sections of jejunum tissue from control (a, c) and infected (b, d) farm piglets four days after inoculation were stained with haematoxylin and eosin (a, b) or rabbit anti-SADSr-CoV N serum (red), DAPI (blue) and mouse antibodies against epithelial cell markers cytokeratin 8, 18 and 19 (green) in (c, d). SADS-CoV N protein is evident in epithelial cells and deeper in the tissue of infected piglets, which exhibit villus shortening. Scale bars, 200 μm (a, b) and 50 μm (c, d). The experiment was conducted three times independently with similar results. The current study highlights the value of proactive viral discovery in wildlife, and targeted surveillance in response to an emerging infectious disease event, as well as the disproportionate importance of bats as reservoirs of viruses that threaten veterinary and public health1. It also demonstrates that by using modern technological platforms, such as NGS, luciferase immunoprecipitation system serology and phylogenetic analysis, key experiments that traditionally rely on the isolation of live virus can be performed rapidly before virus isolation. Bats were captured and sampled in their natural habitat in Guangdong province (Extended Data Fig. 1) as described previously4. Faecal swab samples were collected in viral transport medium (VTM) composed of Hank’s balanced salt solution at pH 7.4 containing BSA (1%), amphotericin (15 μg ml−1), penicillin G (100 units ml−1) and streptomycin (50 μg ml−1). Stool samples from sick pigs were collected in VTM. When appropriate and feasible, intestinal samples were also taken from deceased animals. Samples were aliquoted and stored at –80 °C until use. Blood samples were collected from recovered sows and workers on the farms who had close contact with sick pigs. Serum was separated by centrifugation at 3,000g for 15 min within 24 h of collection and preserved at 4 °C. Human serum collection was approved by the Medical Ethics Committee of the Wuhan School of Public Health, Wuhan University and Hummingbird IRB. Human, pigs and bats were sampled without gender or age preference unless indicated (for example, piglets or sows). No statistical methods were used to predetermine sample size. The following cells were used for virus isolation in this study: Vero (cultured in DMEM and 10% FBS); Rhinolophus sinicus primary or immortalized cells generated in our laboratory (all cultured in DMEM/F12 and 15% FBS): kidney primary cells (RsKi9409), lung primary cells (RsLu4323), lung immortalized cells (RsLuT), brain immortalized cells (RsBrT) and heart immortalized cells (RsHeT); and swine cell lines: two intestinal porcine enterocytes cell lines, IPEC (RPMI1640 and 10% FBS) and SIEC (DMEM and 10% FBS), three kidney cell lines PK15, LLC-PK1 (DMEM and 10% FBS for both) and IBRS (MEM and 10% FBS), and one pig testes cell line, ST (DMEM and 10% FBS). All cell lines were tested free of mycoplasma contamination, species were confirmed and authenticated by microscopic morphologic evaluation. None of the cell lines was on the list of commonly misidentified cell lines (by the ICLAC). Cultured cell monolayers were maintained in their respective medium. PCR-positive pig faecal samples or the supernatant from homogenized pig intestine (in 200 μl VTM) were spun at 8,000g for 15 min, filtered and diluted 1:2 with DMEM supplemented with 16 μg ml−1 trypsin before addition to the cells. After incubation at 37 °C for 1 h, the inoculum was removed and replaced with fresh culture medium containing antibiotics (below) and 16 μg ml−1 trypsin. The cells were incubated at 37 °C and observed daily for cytopathic effect (CPE). Four blind passages (three-day interval between every passage) were performed for each sample. After each passage, both the culture supernatant and cell pellet were examined for the presence of virus by RT–PCR using the SADS-CoV primers listed in Supplementary Table 2. Penicillin (100 units ml−1) and streptomycin (15 μg ml−1) were included in all tissue culture media. Whenever commercial kits were used, the manufacturer’s instructions were followed without modification. RNA was extracted from 200 μl of swab samples (bat), faeces or homogenized intestine (pig) with the High Pure Viral RNA Kit (Roche). RNA was eluted in 50 μl of elution buffer and used as the template for RT–PCR. Reverse transcription was performed using the SuperScript III kit (Thermo Fisher Scientific). To amplify S1 genes from bat samples, nested PCR was performed with primers designed based on HKU2-CoV (GenBank accession number NC_009988.1)19 (Supplementary Table 2). The 25-μl first-round PCR mixture contained 2.5 μl 10× PCR reaction buffer, 5 pmol of each primer, 50 mM MgCl2, 0.5 mM dNTP, 0.1 μl Platinum Taq Enzyme (Thermo Fisher Scientific) and 1 μl cDNA. The 50-μl second-round PCR mixture was identical to the first-round PCR mixture except for the primers. Amplification of both rounds was performed as follows: 94 °C for 5 min followed by 60 cycles at 94 °C for 30 s, 50 °C for 40 s, 72 °C for 2.5 min, and a final extension at 72 °C for 10 min. PCR products were gel-purified and sequenced. For qPCR analysis, primers based on SADS-CoV RdRp and N genes were used (Supplementary Table 2). RNA extracted from above was reverse-transcribed using PrimeScript RT Master Mix (Takara). The 10 μl qPCR reaction mix contained 5 μl 2× SYBR premix Ex TaqII (Takara), 0.4 μM of each primer and 1 μl cDNA. Amplification was performed as follows: 95 °C for 30 s followed by 40 cycles at 95 °C for 5 s, 60 °C for 30 s, and a melting curve step. The SADS-CoV S1 gene was codon-optimized for eukaryotic expression, synthesized (GenScript) and cloned in frame with the Renilla luciferase gene (Rluc) and a Flag tag in the pREN2 vector21. pREN2-S1 plasmids were transfected into Cos-1 cells using Lipofectamine 2000 (Thermo Fisher Scientific). At 48 h post-transfection, cells were collected, lysed and a luciferase assay was performed to determine Rluc expression for both the empty vector (pREN2) and the pREN2-S1 construct. For testing of unknown pig or human serum samples, 1 μl of serum was incubated with 10 million units of Rluc alone (vector) or Rluc-S1, respectively, together with 3.5 μl of a 30% protein A/G UltraLink resin suspension (Pierce, Thermo Fisher Scientific). After extensive washing to remove unbounded luciferase-tagged antigens, the captured luciferase amount was determined using the commercial luciferase substrate kit (Promega). The ratio of Rluc-S1:Rluc (vector) was used to determine the specific S1 reactivity of pig and human sera. Commercial Flag antibody (Thermo Fisher Scientific) was used as the positive control, and various pig sera (from uninfected animals in China or Singapore; or pigs infected with PEDV, TGEV or Nipah virus) were used as a negative control. The N gene from SADSr-CoV 3755 (GenBank accession number MF094702), which shares a 98% amino acid sequence identity to the SADS-CoV N protein, was inserted into pET-28a+ (Novagen) for prokaryotic expression. Transformed Escherichia coli were grown at 37 °C for 12–18 h in medium containing 1 mM IPTG. Bacteria were collected by centrifugation and resuspended in 30 ml of 5 mM imidazole and lysed by sonication. The lysate, from which N protein expression was confirmed with an anti-His-tag antibody, was applied to Ni2+ resin (Thermo Fisher Scientific). The purified N protein, at a concentration of 400 μg ml−1, was used to immunize rabbits for antibody production following published methods27. After immunization and two boosts, rabbits were euthanized and sera were collected. Rabbit anti-N protein serum was used 1:10,000 for subsequent western blots. Construction of expression clones for human ACE2 in pcDNA3.1 has been described previously5, 28. Human DPP4 was amplified from human cell lines. Human APN (also known as ANPEP) was commercially synthesized. Swine APN (also known as ANPEP), DPP4 and ACE2 were amplified from piglet intestine. Full-length gene fragments were amplified using specific primers (provided upon request). Human ACE2 was cloned into pCDNA3.1 fused with a His tag. Human APN and DPP4, swine APN, DPP4 and ACE2 were cloned into pCAGGS fused with an S tag. Purified plasmids were transfected into HeLa cells. After 24 h, expression human or swine genes in HeLa cells was confirmed by immunofluorescence assay using mouse anti-His tag or mouse anti-S tag monoclonal antibodies (produced in house) followed by Cy3-labelled goat anti-mouse/rabbit IgG (Proteintech Group). The codon-humanized S genes of SADS-CoV or MERS-CoV cloned into pcDNA3.1 were used for pseudovirus construction as described previously5, 28. In brief, 15 μg of each pHIV-Luc plasmid (pNL4.3.Luc.R-E-Luc) and the S-protein-expressing plasmid (or empty vector control) were co-transfected into 4 × 106 HEK293T cells using Lipofectamine 3000 (Thermo Fisher Scientific). After 4 h, the medium was replaced with fresh medium. Supernatants were collected 48 h after transfection and clarified by centrifugation at 3,000g, then passed through a 0.45-μm filter (Millipore). The filtered supernatants were stored at −80 °C in aliquots until use. To evaluate the incorporation of S proteins into the core of HIV virions, pseudoviruses in supernatant (20 ml) were concentrated by ultracentrifugation through a 20% sucrose cushion (5 ml) at 80,000g for 90 min using a SW41 rotor (Beckman). Pelleted pseudoviruses were dissolved in 50 μl phosphate-buffered saline (PBS) and examined by electron microscopy. HeLa cells transiently expressing APN, ACE2 or DPP4 were prepared using Lipofectamine 2000 (Thermo Fisher Scientific). Pseudoviruses prepared above were added to HeLa cells overexpressing APN, ACE2 or DPP4 24 h after transfection. The unabsorbed viruses were removed and replaced with fresh medium at 3 h after infection. The infection was monitored by measuring the luciferase activity conferred by the reporter gene carried by the pseudovirus, using the Luciferase Assay System (Promega) as follows: cells were lysed 48 h after infection, and 20 μl of the lysates was taken for determining luciferase activity after the addition of 50 μl of luciferase substrate. HeLa cells transiently expressing APN, ACE2 or DPP4 were prepared using Lipofectamine 2000 (Thermo Fisher Scientific) in a 96-well plate, with mock-transfected cells as controls. SADS-CoV grown in Vero cells was used to infect HeLa cells transiently expressing APN, ACE2 or DPP4. The inoculum was removed after 1 h of absorption and washed twice with PBS and supplemented with medium. SARS-related-CoV WIV167 and MERS-CoV HIV-pseudovirus were used as positive control for human/swine ACE2 or human/swine DPP4, respectively. After 24 h of infection, cells were washed with PBS and fixed with 4% formaldehyde in PBS (pH 7.4) for 20 min at room temperature. SARS-related-CoV WIV16 replication was detected using rabbit antibody against the SARS-related-CoV Rp3 N protein (made in house, 1:100) followed by Cy3-conjugated goat anti-rabbit IgG (1:50, Proteintech)7. SADS-CoV replication was monitored using rabbit antibody against the SADSr-CoV 3755 N protein (made in house, 1:50) followed by FITC-conjugated goat anti-rabbit IgG (1:50, Proteintech). Nuclei were stained with DAPI (Beyotime). Staining patterns were examined using confocal microscopy on a FV1200 microscope (Olympus). Infection of MERS-CoV HIV-pseudovirus was monitored by luciferase 48 h after infection. Tissue from the small intestine of deceased pigs was homogenized and filtered through 0.45-μm filters before nucleic acid extraction and ribosomal RNA was depleted using the NEBNext rRNA Depletion Kit (New England Biolabs). Metagenomics analysis of both RNA and DNA viruses was performed. For RNA virus screening, the sequencing library was constructed using Ion Total RNA-Seq Kit v2 (Thermo Fisher Scientific). For DNA virus screening, NEBNext Fast DNA Fragmentation & Library Prep Set for Ion Torrent (New England Biolabs) was used for library preparation. Both libraries were sequenced on an Ion S5 sequencer (Thermo Fisher Scientific). An analysis pipeline was applied to the sequencing data, which included the following analysis steps: (1) raw data quality filtering; (2) host genomic sequence filtering; (3) BLASTn search against the virus nucleotide database using BLAST; (4) BLASTx search against the virus protein database using DIAMOND v.0.9.0; (5) contig assembling and BLASTx search against the virus protein database. For whole viral genome sequencing, amplicon primers (provided upon request) were designed using the Thermo Fisher Scientific online tool with the HKU2-CoV and the SADS-CoV farm A genomes as references, and the sequencing libraries were constructed using NEBNext Ultra II DNA Library Prep Kit for Illumina and sequenced on an MiSeq sequencer. PCR and Sanger sequencing was performed to fill gaps in the genome. Genome sequences were assembled using CLC Genomic Workbench v.9.0. 5′-RACE was performed to determine the 5′-end of the genomes using SMARTer RACE 5′/3′ Kit (Takara). Genomes were annotated using Clone Manager Professional Suite 8 (Sci-Ed Software). SADS-CoV genome sequences and other representative coronavirus sequences (obtained from GenBank) were aligned using MAFFT v.7.221. Phylogenetic analyses with full-length genome, S gene and RdRp were performed using MrBayes v.3.2. Markov chain Monte Carlo was run for 20–50 million steps using the GTR+G+I model (general time reversible model of nucleotide substitution with a proportion of invariant sites and γ-distributed rates among sites). The first 10% was removed as burn-in. The association between phylogenies and phenotypes (for example, host species and farms) was assessed by BaTS beta-build2, with the trees obtained in the previous step used as input. For SADS-CoVs, a median-joining network analysis was performed using PopART v.1.7, with ɛ = 0. Phylogenetic analysis of the 33 full-length SADS-CoV genome sequences was performed using RAxML v.8.2.11, with GTRGAMMA as the nucleotide substitution model and 1,000 bootstrap replicates. The maximum likelihood tree was used to test the molecular clock using TempEst v.1.5. Potential genetic recombination events in our datasets were detected using RDP v.4.72. Experiments were carried out strictly in accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The use of animals in this study was approved by the South China Agricultural University Committee of Animal Experiments (approval number 201004152). Two different animal challenge experiments were conducted. Pigs were used without gender preference. In the first experiment, which was conducted before the virus was isolated, we used three-day old specific pathogen-free (SPF) piglets of the same breeding line, cared for at a SPF facility, fed with colostrum (except one). These piglets were bred and reared to be free of PEDV, CSFV, SIV, PCV2 and PPV infections, and were routinely tested for viral infections using PCR. We also conducted NGS to further confirm that these were animals were free of infection of the above viruses before the animal experiment, and to demonstrate that the animals were free of SADS-CoV infection. The intestinal tissue samples from healthy and diseased animals (intestinal samples excised from euthanized piglets, then ground to make slurry for the inoculum and NGS was performed to confirm no other pig pathogens were found in the samples), were used to feed two groups of 5 (control) and 7 (infection) animals, respectively. For the second experiment, isolated SADS-CoV was used to infect healthy piglets from a farm in Guangdong, which had been free of diarrheal disease for a number of weeks. These piglets were from the same breed as those on SADS-affected farms, to eliminate potential host factor differences and to more accurately reproduce the conditions that occurred during the outbreak in the region. Both groups of piglets were cared for at a known pig disease-free facility. Again, qPCR and NGS were used to make sure that there was no other known swine diarrhoea virus present in the virus inoculum or any of the experimental animals. Two groups (6 for each group) of three-day old piglets were inoculated with SADS-CoV culture supernatant or normal cell culture medium as control. NGS and qPCR were used to confirm that there were no other known swine pathogens in the inoculum. For both experiments, animals were recorded daily for signs of diseases, such as diarrhoea, weight loss and death. Faecal swabs were collected daily from all animals and screened for known swine diarrhoea viruses by qPCR. Weight loss was calculated as the percentage weight loss compared the original weight at day 0 with a threshold of >5%. It is important to point out that piglets when they are three days old tend to suffer from diarrhoea and weight loss when they are taken away from sows and the natural breast-feeding environment even without infection. At experimental endpoints, piglets were humanely euthanized and necropsies performed. Pictures were taken to record gross pathological changes to the intestines. Ileal, jejunal and duodenal tissues were taken from selected animals and stored at –80 °C for further analysis. Frozen (–80 °C) small intestinal tissues including duodenum, jejunum and ileum taken from the experimentally infected pigs were pre-frozen at –20 °C for 10 min. Tissues were then embedded in optimal cutting temperature (OCT) compound and cut into 8-μm sections using the Cryotome FSE machine (Thermo Fisher Scientific). Mounted microscope slides were fixed with paraformaldehyde and stained with haematoxylin and eosin for histopathological examination. For immunohistochemistry analysis, a rabbit antibody raised against the SADSr-CoV 3755 N protein was used for specific staining of SADS-CoV antigen. Slides were blocked by incubating with 10% goat serum (Beyotime) at 37 °C for 30 min, followed by overnight incubation at 4 °C with the rabbit anti-3755 N protein serum (1:1,000) and mouse anti-cytokeratin 8+18+19 monoclonal antibody (Abcam), diluted 1:100 in PBST buffer containing 5% goat serum. After washing, slides were then incubated for 50 min at room temperature with Cy3-conjugated goat-anti-rabbit IgG (Proteintech) and FITC-conjugated goat-anti-mouse IgG (Proteintech), diluted 1:100 in PBST buffer containing 5% goat serum. Slides were stained with DAPI (Beyotime) and observed under a fluorescence microscope (Nikon). Further information on experimental design is available in the Nature Research Reporting Summary linked to this paper. Sequence data that support the findings of this study have been deposited in GenBank with accession codes MF094681–MF094688, MF769416–MF769444, MF094697–MF094701, MF769406–MF769415 and MG557844. Raw sequencing data that support the findings of this study have been deposited in the Sequence Read Achieve (SRA) with accession codes SRR5991648, SRR5991649, SRR5991650, SRR5991651, SRR5991652, SRR5991654, SRR5991655, SRR5991656, SRR5991657, SRR5991658 and SRR5995595. The Extended Data Figures and Tables section originally published with this article was missing Tables 1–5. This has now been corrected. 1. Olival, K. J. et al. Host and viral traits predict zoonotic spillover from mammals. Nature 546, 646–650 (2017). 2. Guan, Y. et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 302, 276–278 (2003). 3. Lau, S. K. et al. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc. Natl Acad. Sci. USA 102, 14040–14045 (2005). 4. Li, W. et al. Bats are natural reservoirs of SARS-like coronaviruses. Science 310, 676–679 (2005). 5. Ge, X. Y. et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature 503, 535–538 (2013). 6. He, B. et al. Identification of diverse alphacoronaviruses and genomic characterization of a novel severe acute respiratory syndrome-like coronavirus from bats in China. J. Virol. 88, 7070–7082 (2014). 7. Yang, X. L. et al. Isolation and characterization of a novel bat coronavirus closely related to the direct progenitor of severe acute respiratory syndrome coronavirus. J. Virol. 90, 3253–3256 (2016). 8. Wu, Z. et al. ORF8-related genetic evidence for Chinese horseshoe bats as the source of human severe acute respiratory syndrome coronavirus. J. Infect. Dis. 213, 579–583 (2016). 9. Wang, L. et al. Discovery and genetic analysis of novel coronaviruses in least horseshoe bats in southwestern China. Emerg. Microbes Infect. 6, e14 (2017). 10. Hu, B. et al. Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus. PLoS Pathog. 13, e1006698 (2017). 11. Drosten, C. et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Engl. J. Med. 348, 1967–1976 (2003). 12. Ksiazek, T. G. et al. A novel coronavirus associated with severe acute respiratory syndrome. N. Engl. J. Med. 348, 1953–1966 (2003). 13. Marra, M. A. et al. The genome sequence of the SARS-associated coronavirus. Science 300, 1399–1404 (2003). 14. Peiris, J. S. et al. Coronavirus as a possible cause of severe acute respiratory syndrome. Lancet 361, 1319–1325 (2003). 15. Rota, P. A. et al. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science 300, 1394–1399 (2003). 16. Wang, L.-F. & Cowled, C. (eds) Bats and Viruses: A New Frontier of Emerging Infectious Diseases 1st edn (John Wiley & Sons, Hoboken, 2015). 17. Dong, N. et al. Porcine deltacoronavirus in mainland China. Emerg. Infect. Dis. 21, 2254–2255 (2015). 18. Sun, D., Wang, X., Wei, S., Chen, J. & Feng, L. Epidemiology and vaccine of porcine epidemic diarrhea virus in China: a mini-review. J. Vet. Med. Sci. 78, 355–363 (2016). 19. Lau, S. K. et al. Complete genome sequence of bat coronavirus HKU2 from Chinese horseshoe bats revealed a much smaller spike gene with a different evolutionary lineage from the rest of the genome. Virology 367, 428–439 (2007). 20. Chen, J. et al. Molecular epidemiology of porcine epidemic diarrhea virus in China. Arch. Virol. 155, 1471–1476 (2010). 21. Burbelo, P. D. et al. Serological diagnosis of human herpes simplex virus type 1 and 2 infections by luciferase immunoprecipitation system assay. Clin. Vaccine Immunol. 16, 366–371 (2009). 22. Gong, L. et al. A new bat-HKU2-like coronavirus in swine, China, 2017. Emerg. Infect. Dis. 23, 1607–1609 (2017). 23. Pan, Y. et al. Discovery of a novel swine enteric alphacoronavirus (SeACoV) in southern China. Vet. Microbiol. 211, 15–21 (2017). 24. Li, W. et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 426, 450–454 (2003). 25. Masters, P. S. & Perlman, S. in Fields Virology Vol. 2 (eds Knipe, D. M. & Howley, P. M.) 825–858 (Lippincott Williams & Wilkins, Philadelphia, 2013). 26. Raj, V. S. et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 495, 251–254 (2013). 27. Harlow, E. & Lane, D. Antibodies: A Laboratory Manual (Cold Spring Harbor Laboratory Press, New York, 1988). 28. Ren, W. et al. Difference in receptor usage between severe acute respiratory syndrome (SARS) coronavirus and SARS-like coronavirus of bat origin. J. Virol. 82, 1899–1907 (2008). We thank S.-B. Xiao for providing pig cell lines, P. Burbelo for providing the luciferase immunoprecipitation system vector and L. Zhu for enabling the rapid synthesis of the S gene; the WIV animal facilities; J. Min for help with the preparation of the immunohistochemistry samples; and G.-J. Zhu and A. A. Chmura for assistance with bat sampling. This work was jointly supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (XDPB0301) to Z.-L.S., China Natural Science Foundation (81290341 and 31621061 to Z.-L.S., 81661148058 to P.Z., 31672564 and 31472217 to J.-Y.M., 81572045, 81672001 and 81621005 to Y.-G.T.), National Key Research and Development Program of China (2015AA020108, 2016YFC1202705, AWS16J020 and AWS15J006) to Y.-G.T.; National Science and Technology Spark Program (2012GA780026) and Guangdong Province Agricultural Industry Technology System Project (2016LM1112) to J.-Y.M., State Key Laboratory of Pathogen and Biosecurity (SKLPBS1518) to Y.-G.T., Taishan Scholars program of Shandong province (ts201511056 to W.-F.S.), NRF grants NRF2012NRF-CRP001–056, NRF2016NRF-NSFC002-013 and NMRC grant CDPHRG/0006/2014 to L.-F.W., Funds for Environment Construction & Capacity Building of GDAS’ Research Platform (2016GDASPT-0215) to LBZ, United States Agency for International Development Emerging Pandemic Threats PREDICT project (AID-OAA-A-14-00102), National Institute of Allergy and Infectious Diseases of the National Institutes of Health (Award Number R01AI110964) to P.D. and Z.-L.S. Nature thanks C. Drosten, G. Palacios and L. Saif for their contribution to the peer review of this work. L.-F.W., Z.-L.S., P.Z., Y.-G.T., and J.-Y.M. conceived the study. P.Z., W.Z., Y.Z., S.M., X.-S.Z., B.L., X.-L.Y., H.G., D.E.A., Y.L., X.L.L. and J.C. performed qPCR, serology and histology experiments and cultured the virus. H.F., Y.-W.Z., J.-M.L., G.-Q.P., X.-P.A., Z.-Q.M., T.-T.H., Y.H., Q.S., Y.-Y.W., S.-Z.X., X.-L.-L.Z., W.-F.S. and J.L. performed genome sequencing and annotations. T.L., Q.-M.X., J.-W.C., L.Z., K.-J.M., Z.-X.W., Y.-S.C., D.L., Y.S., F.C., P.-J.G. and R.H. prepared the samples and carried out animal challenge experiments. Z.-L.S., P.D., L.-B.Z., S.-Y.L. coordinated collection of bat samples. P.Z., L.-F.W., Z.-L.S. and P.D. had a major role in the preparation of the manuscript. Correspondence to Peter Daszak or Lin-Fa Wang or Zheng-Li Shi or Yi-Gang Tong or Jing-Yun Ma. The authors declare no competing interests. Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. a, SADS-affected farms are labelled (farms A–D) with blue swine silhouettes following the temporal sequence of the outbreaks. Bat sampling sites are indicated with black bat silhouettes. The bat SADSr-CoV that is most closely related to SADS-CoV (sample 162140) originated in Conghua. The red flag marks Foshan city, the site of the SARS index case. b, Pooled intestinal samples (n = 5 or more biological independent samples) were collected at dates given on the x axis from deceased piglets and analysed by qPCR. The viral load for each piglet is shown as copy number per milligram of intestine tissue (y axis). a, Bayesian phylogenetic tree of the full-length genome. b, Bayesian phylogenetic tree of the ORF1a and ORF1b sequences. Trees were constructed using MrBayes with the average standard deviation of split frequencies under 0.01. The host of each sequence is represented as a silhouette. Newly sequenced SADS-CoVs are highlighted in red, bat SADSr-CoVs are shown in blue and previously published sequences are shown in black. Scale bars, nucleotide substitutions per site. a, Phylogenetic tree constructed using MrBayes. The GTR+GAMMA model was applied and 20 million steps were run, with the first 10% removed as burn in. Viruses from different farms are labelled with different colours. Scale bar, nucleotide substitutions per site. b, Median-joining haplotype network constructed using ProART. In this analysis, ɛ = 0 was used. The size of the circles represents the number of samples. The larger the circle, the more samples it includes. The potential genetic recombination events were detected using RDP. For each virus strain, different colours represent different sources of the genomes. Source data a, b, Vero cells are shown 20 h after infection with mock (a) or SADS-CoV (b). c, d, Mock or SADS-CoV-infected samples stained with rabbit serum raised against the recombinant SADSr-CoV N protein (red) and DAPI (blue). The experiment was conducted independently three times with similar results. Scale bars, 100 μm. Source data Extended Data Table 1 List of all known swine viruses tested by PCR at the beginning of the of SADS outbreak investigation on the four farms Extended Data Table 2 List of SADSr-CoVs detected in bats in Guangdong, China Extended Data Table 3 Test of SADS-CoV entry and infection in Hela cells expressing known coronavirus receptors Extended Data Table 4 Experimental infection of SPF piglets using intestine tissue homogenate Extended Data Table 5 Experimental animal infection of farm piglets using cultured SADS-CoV This file contains two supplementary tables. Supplementary table 1 contains a list of nucleotide variants among the 33 SADS-CoV genomes obtained from the four farms. Supplementary table 2 contains a list of PCR primers used in this study Zhou, P., Fan, H., Lan, T. et al. Fatal swine acute diarrhoea syndrome caused by an HKU2-related coronavirus of bat origin. Nature 556, 255–258 (2018). https://doi.org/10.1038/s41586-018-0010-9 Received07 July 2017 Accepted26 February 2018 Published04 April 2018 Issue Date12 April 2018 Get shareable link The Lancet (2020) Transboundary and Emerging Diseases (2020) Virology (2020) The Lancet (2020) Biosafety and Health (2020)
The discovery of SARS-like coronavirus in bats suggests that bats could be the natural reservoir of SARS-CoV. However, previous studies indicated the angiotensin-converting enzyme 2 (ACE2) protein, a known SARS-CoV receptor, from a horseshoe bat was unable to act as a functional receptor for SARS-CoV. Here, we extended our previous study to ACE2 molecules from seven additional bat species and tested their interactions with human SARS-CoV spike protein using both HIV-based pseudotype and live SARS-CoV infection assays. The results show that ACE2s of Myotis daubentoni and Rhinolophus sinicus support viral entry mediated by the SARS-CoV S protein, albeit with different efficiency in comparison to that of the human ACE2. Further, the alteration of several key residues either decreased or enhanced bat ACE2 receptor efficiency, as predicted from a structural modeling study of the different bat ACE2 molecules. These data suggest that M. daubentoni and R. sinicus are likely to be susceptible to SARS-CoV and may be candidates as the natural host of the SARS-CoV progenitor viruses. Furthermore, our current study also demonstrates that the genetic diversity of ACE2 among bats is greater than that observed among known SARS-CoV susceptible mammals, highlighting the possibility that there are many more uncharacterized bat species that can act as a reservoir of SARS-CoV or its progenitor viruses. This calls for continuation and expansion of field surveillance studies among different bat populations to eventually identify the true natural reservoir of SARS-CoV. Severe acute respiratory syndrome coronavirus (SARS-CoV) is the aetiological agent responsible for the SARS outbreaks during 2002–2003, which had a huge global impact on public health, travel and the world economy [4, 11]. The host range of SARS-CoV is largely determined by the specific and high-affinity interactions between a defined receptor-binding domain (RBD) on the SARS-CoV spike protein and its host receptor, angiontensin-converting enzyme 2 (ACE2) [6, 7, 9]. It has been hypothesized that SARS-CoV was harbored in its natural reservoir, bats, and was transmitted directly or indirectly from bats to palm civets and then to humans [10]. However, although the genetically related SARS-like coronavirus (SL-CoV) has been identified in horseshoe bats of the genus Rhinolophus [5, 8, 12, 18], its spike protein was not able to use the human ACE2 (hACE2) protein as a receptor [13]. Close examination of the crystal structure of human SARS-CoV RBD complexed with hACE2 suggests that truncations in the receptor-binding motif (RBM) region of SL-CoV spike protein abolish its hACE2-binding ability [7, 10], and hence the SL-CoV found recently in horseshoe bats is unlikely to be the direct ancestor of human SARS-CoV. Also, it has been shown that the human SARS-CoV spike protein and its closely related civet SARS-CoV spike protein were not able to use a horseshoe bat (R. pearsoni) ACE2 as a receptor [13], highlighting a critical missing link in the bat-to-civet/human transmission chain of SARS-CoV. There are at least three plausible scenarios to explain the origin of SARS-CoV. First, some unknown intermediate hosts were responsible for the adaptation and transmission of SARS-CoV from bats to civets or humans. This is the most popular theory of SARS-CoV transmission at the present time [10]. Second, there is an SL-CoV with a very close relationship to the outbreak SARS-CoV strains in a non-bat animal host that is capable of direct transmission from reservoir host to human or civet. Third, ACE2 from yet to be identified bat species may function as an efficient receptor, and these bats could be the direct reservoir of human or civet SARS-CoV. Unraveling which scenario is most likely to have occurred during the 2002–2003 SARS epidemic is critical for our understanding of the dynamics of the outbreak and will play a key role in helping us to prevent future outbreaks. To this end, we have extended our studies to include ACE2 molecules from different bat species and examined their interaction with the human SARS-CoV spike protein. Our results show that there is great genetic diversity among bat ACE2 molecules, especially at the key residues known to be important for interacting with the viral spike protein, and that ACE2s of Myotis daubentoni and Rhinolophus sinicus from Hubei province can support viral entry. HeLa cells were grown in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (Gibco, USA). Rabbit polyclonal antibodies against ACE2 of R. pearsoni (RpACE2) were generated using R. pearsoni ACE2 protein expressed in Escherichia coli at the Wuhan Institute of Virology following standard procedures. Bats were sampled from their natural habitats in Hubei, Guangxi, Guizhou, Henan and Yunnan provinces in China as described previously [8]. Bat identification was initially determined in the field by morphology and later confirmed in the laboratory by sequencing the mitochondrial cytochrome b gene from samples of blood cells or rectal tissue as described previously [1]. Total RNA was extracted from bat rectal tissue using TRIzol Reagent (Invitrogen, USA) and treating with RNase-free DNase I at 37°C for 30 min. First-strand cDNA was synthesized from total RNA by reverse transcription with random hexamers. Full-length bat ACE2 fragments were amplified using the forward primer bAF2 (5′-CTTGGTACCATGTCAGGCTCTTYCTGG-3′) and the reverse primer bAR2 (5′-CCGCTCGAGCTAAAAB[G/T/C]GAV[G/A/C]GTCTGAACATCATC-3′). The PCR mixture (25 μL) contained 0.5 μL cDNA, 1.5 mM MgCl2 and 0.2 μM of each primer, and the fragments were amplified using the following parameters: 95°C for 5 min, 35 cycles of 94°C for 30 s, 55°C for 45 s and 68°C for 3 min, with a final elongation step at 68°C for 10 min. All bat ACE2s were cloned into pCDNA3.1 with KpnI and XhoI, and this was verified by sequencing. For samples in which full-length ACE2 amplification was unsuccessful, the N-terminal region (1–1,170 bp) was amplified using the forward primer bAF2 and the reverse primer RMR (5′-TTAGCTCCATTTCTTAGCAGGTAGG-3′). Chimeric ACE2 was constructed by combining the N-terminal region of bat ACE2 with the C-terminal portion of human ACE2 at the unique BamHI site (1,070–1,075 bp). The chimera was subsequently cloned into pCDNA3.1 with KpnI and XhoI and sequenced as above. ACE2 from M. daubentoni was chosen to generate a series of ACE2 mutants using a QuikChange II Site-Directed Mutagenesis Kit (Stratagene, USA). The altered amino acid codon for each mutant is indicated as follows: I27T, N31K, K35E, and H41Y. Mutants were confirmed by sequencing. All bat ACE2s were submitted to GenBank (EF569964, GQ999931–GQ999938). Sequence alignment was performed using ClustalX version 1.83 [15] and corrected manually. A phylogenetic tree based on amino acid (aa) sequences was constructed using the neighbor-joining (NJ) method in MEGA version 4.1. [14]. Lysates of HeLa cells expressing human ACE2 or bat ACE2 were separated on a 4–10% SDS-PAGE gel, followed by transfer to a polyvinylidene difluoride (PVDF) membrane using a semi-dry protein transfer apparatus (Bio-Rad, USA). The membrane was probed with a rabbit polyclonal antibody against the bat ACE2 protein (1:200) at room temperature for 1 h, followed by incubation with alkaline-phosphatase-conjugated goat anti-rabbit IgG (1:1,000) (Chemicon, Australia). The probed proteins were visualized using NBT and BCIP color development (Promega, USA). An HIV-1-luciferase pseudotype virus carrying the SARS-CoV BJ01 S protein, HIV/BJ01-S, was prepared as described previously [13]. HeLa cells were seeded onto 96-well plates for 18 h and then transfected with 0.2 μg recombinant plasmid containing bat or human ACE2 using 0.5 μL Lipofectamine 2000 (Invitrogen, USA) according to the manufacturer’s protocol. At 24 h post-transfection, 30 μL medium containing HIV/BJ01-S was added to each well. At 2–3 h postinfection, unadsorbed viruses were removed, and fresh medium was added. The infection was monitored by measuring luciferase activity, expressed from the reporter gene carried by the pseudovirus, using a luciferase assay kit (Promega, USA). Cells were lysed at 48 h postinfection by adding 20 μL lysis buffer provided with the kit, and 10 μL of the resulting lysates were tested for luciferase activity by the addition of 20 μL luciferase substrate in a Turner Designs TD-20/20 luminometer. Each infection experiment was conducted in triplicate, and all experiments were repeated three times. Live SARS-CoV infection was carried out under BSL4 conditions at the Australian Animal Health Laboratory (AAHL) as described previously [16, 17]. Briefly, 48 h after transfection, the time at which expression of the ACE2 receptor on the HeLa cell surface is optimal, 2 × 106 TCID50 of virus was added to the cells for infection. The cells were fixed 24 h later by treatment with 100% methanol for 10 min and washed five times with PBST. The primary antibody, chicken anti-SARS-CoV S (produced against the recombinant S protein expressed in E. coli at AAHL), at a 1:500 dilution in 1% BSA/PBS, was added and incubated with the cells for 1 h at room temperature. An FITC anti-chicken conjugate (Chemicon, Australia) at 1:1,000 in 1% BSA/PBS was added after washing the cells five times and incubated with the cells for 1 h. Infection was monitored by immunofluorescent microscopic analysis. ACE2 genes from seven bat species were amplified and cloned (Fig. 1, sFig. 1). Full-length genes were obtained from Rhinolophus ferrumequinum from Hubei province (Rf-HB), R. macrotis from Hubei province (Rm-HB), R. pearsoni from Guangxi (Rp-GX), R. pusillus from Hubei province (Rpu-HB), R. sinicus from Guangxi province (Rs-GX) and R. sinicus from Hubei province (Rs-HB). For the following bat species, amplification of the full-length coding region was not successful, and instead,the N-terminal region was cloned in frame with the C-terminal region of the human ACE2 gene to form a chimeric full-length ACE molecule: R. pearsoni from Guizhou province (Rp-GZ), Myotis daubentonii bat from Yunnan province (Md-YN) and Hipposideros pratti bat from Henan province (Hp-HN). The full-length sequences of bat ACE2 are identical in size to that of hACE2 (805 aa in total). Sequence comparison showed that bat ACE2s are closely related to ACE2s of other mammals and have an aa sequence identity of 80–82% to human and civet ACE2. The aa identity of ACE2 from different bat families ranges from 78 to 84%, and within the genus Rhinolophus, the sequence identity increases to 89–98%. The major sequence variation among bat ACE2s is located in the N-terminal region, which has been identified in structural studies as the SARS-CoV-binding region [6, 7]. A phylogenetic tree was constructed based on the sequences of bat ACE2 (sFig. 2) using the MEGA package [14]. Sequence alignment of SARS-CoV binding regions of ACE2s from 9 bats, civet and human. The GenBank accession numbers of bat, civet and human ACE2 are as follows: human (NM021804), civet (AY881174), Rf-HB (GQ999931), Rm-HB (GQ999932), Rs-GX (GQ999933), Rp-GX (EF569964), Hp-HN (GQ999934), Rp-GZ (GQ999935), Rs-HB (GQ999936), Md-YN(GQ999937) and Rpu-HB (GQ999938). The alignment was generated using ClustalX v1.83. In black are single, fully conserved residues. In gray are strongly conserved residues. In light gray are weakly conserved residues. Asterisks indicate residues that interact directly with the receptor-binding domain of the SARS-CoV S protein All ACE2 genes were cloned into a eukaryotic expression vector and used to transfect HeLa cells. Western blot analysis showed that all ACE2s were expressed efficiently and at very similar levels and were recognized by a rabbit anti-bat ACE2 antibody with an apparent molecular weight of approximately 100–130 kDa (Fig. 2c). Testing of the ability of bat ACE2 proteins to mediate pseudovirus HIV/BJ01-S and live SARS- CoV infection. a HeLa cells transfected with plasmids encoding bat and human ACE2s were infected with pseudovirus HIV/BJ01-S. Infectivity was determined by measuring the activity of reporter luciferase as described in “Materials and methods”. HeLa cells transfected with pcDNA3.1 and human ACE2 were used as the negative and positive controls, respectively. All tests were performed in triplicate, and the experiments were repeated three times. The error bar represents the calculated standard deviation. I27T, N31K, K35E, and H41Y are mutants of MdACE2 that were made using a QuikChange II Site-Directed Mutagenesis Kit. b SARS-CoV live virus infection using the ACE2s from bats as described in “Materials and methods”. HeLa cells transfected with pcDNA3.1 and human ACE2 were used as the negative and positive controls, respectively. c Expression of bat or human ACE2. Lysates from HeLa cells transfected with plasmid expressing human or bat ACE2 were analyzed by western blot. Rabbit anti-bat ACE2 polyclonal antibody (upper panel) or β-actin monoclonal antibody (lower panel) was used as the primary antibody. Lane 1 vector pcDNA3.1 control; lanes 2–10 bat ACE2 from samples Rf-HB, Rm-HB, Rpu-HB, Hp-HN, Rp-HB, Rp-GZ, Rs-GX, Rs-HB and Md-YN; lanes 11–14 Md-YN ACE2 mutant I27T, N31K, K35E and H41Y; lane 15 human ACE2. The abbreviations of bat species are given in the main text To examine the susceptibility of different bat ACE2 molecules to SARs-CoV entry, the HIV/BJ01-S pseudovirus system was used to infect HeLa cells transiently expressing bat ACE2 or human ACE2 genes. Among the bat ACE2s, only MdACE2 (MdACE2) and Rs-HB ACE2 demonstrated significant pseudovirus infection, as deduced from the significantly higher level of luciferase activity in comparison to background activity in the negative control (Fig. 2a). Although such assays are not to be viewed as an absolute quantification of receptor activity, it is nevertheless worth noting that MdACE2-mediated infection seemed to be more efficient than with Rs-HB ACE2. In the same context, it is clear that the bat ACE2s were less efficient overall than the human ACE2 in this particular assay system. The biological significance of this observation remains to be determined. The functionality of MdACE2 and Rs-HB ACE2 as SARS-CoV entry receptors was further confirmed by infection with live virus. As shown in Fig. 2b, both bat ACE2 proteins could clearly support SARs-CoV infection. No attempt was made to quantify infection efficiency in this study due to difficulties encountered in conducting experiments under BSL4 conditions. Homologous structural modeling of human SARS-CoV RBD complexed with MdACE2 supports MdACE2 as a receptor for human SARS-CoV S protein. The crystal structure of human SARS-CoV RBD complexed with hACE2 shows that two salt bridges at the SARS-CoV-hACE2 interface, between hACE2 Lys31 and Glu35 and between hACE2 Lys353 and hACE2 Glu38, are both buried in a hydrophobic environment and contribute critically to the SARS-CoV-hACE2 interactions (Fig. 3a, c) [7]. Disturbance of the formation of either of these salt bridges weakens SARS-CoV-hACE2 binding. The Lys31-Glu35 salt bridge at the SARS-CoV-hACE2 interface becomes an Asn31-Lys35 hydrogen bond at the SARS-CoV-Md-YNACE2 interface (Fig. 3b), which possibly weakens virus-receptor binding but still is largely compatible with the virus-receptor interface. Thr27 on hACE2 supports the Lys31-Gu35 salt bridge through hydrophobic interactions with Tyr475 (Fig. 3a); Ile27 on MdACE2 supports the Asn31-Lys35 hydrogen bond more efficiently than Thr27 through tighter hydrophobic interactions with Tyr475 (Fig. 3b). Moreover, Tyr41 on hACE2 supports the Lys353-Glu38 salt bridge (Fig. 3c); His41 on MdACE2 supports the same salt bridge less efficiently than Tyr41 (Fig. 3d). Overall, MdACE2 is an efficient receptor for SARS-CoV, despite the fact that its receptor activity is lower than that of hACE2. Homologous structural modeling of SARS-CoV and Md-YN ACE2 (MdACE2) interactions. a Critical salt bridge between hACE2 Lys31 and Glu35 and the hydrophobic residues surrounding it, based on the experimentally determined crystal structure of SARS-CoV RBD complexed with hACE2 (PDB 2AJF). b Homologous structural modeling of the hydrogen bond between MdACE2 Asn31 and Lys35. The modeling was done in the program O [3]. c Critical salt bridge between hACE2 Lys353 and Glu38 and the hydrophobic residues surrounding it, based on the structure of SARS-CoV RBD complexed with hACE2. d Homologous structural modeling of the salt bridge between MdaACE2 Lys353 and SARS-CoV Glu38 and the hydrophobic residues surrounding it. Structural illustrations were prepared using the program Povscript [2] Compared with MdACE2, Rs-HB ACE2 contains Glu31 and Glu35, which are not compatible with each other due to their same negative charges, which disfavor virus-receptor binding. However, Rs-HB ACE2 also contains Thr27 and Tyr41, both of which support SARS-CoV entry by contributing favorably to the hydrophobic interactions at the virus-receptor interface. Thus, Rs-HB is a low-efficiency receptor for SARS-CoV. All of the other bat ACE2 molecules contain combinations of the aforementioned key residues that are completely incompatible with virus–receptor interactions. More specifically, they either contain same-charged residues at the 31 and 35 positions, which repel each other, or contain His41 and Lys27, which disfavor SARS-CoV binding (Fig. 1). In particular, Lys27 on some of these bat ACE2 molecules is incompatible with certain hydrophobic residues, such as Leu443 and Phe460, on SARS-CoV RBD (Fig. 3a, b). Therefore, these bat ACE2 molecules are not receptors for SARS-CoV. To confirm the above homologous structural analysis, we carried out site-directed mutagenesis on MdACE2. Our results show that mutations E31K, K35E, and I27T all dramatically decrease the receptor activity of MdACE2, whereas mutation H41Y greatly increases its receptor activity (Fig. 2a). Therefore, our mutagenesis data further confirmed that key residues in ACE2 determine the receptor activity of MdACE2. Our finding that M. daubentoni and R. sinicus could support SARS-CoV infection has important implications in relation to the origin of SARS-CoV. Since all lines of investigation have indicated that ACE2-binding affinity is among the important determinants for SARS-CoV host range, our data would suggest that M. Daubentonii and R. sinicus have the potential to serve as the direct reservoirs for human SARS-CoV or its highly related civet SARS-CoV. To further investigate the potential of M. Daubentonii and R. sinicus as reservoirs for SARS-CoV, more efforts will have to be directed toward widening the surveillance of bats in these families and in different geographical locations. Another important finding of our current study is the great genetic diversity of bat ACE2 proteins, which is in contrast to the genetically homogenous hACE2 [10]. Sequence variations of bat ACE2, especially in positions that are critical to SARS-CoV binding, such as residues 27, 31, 35, and 41, suggest that, in addition to the Md-YN and Rs-HB ACE2s, there may be many other bats with an ACE2 protein that makes them susceptible to SARS-CoV entry. This again highlights the need for more field surveillance and molecular characterization of different bat ACE2 proteins until the true reservoir host of SARS-CoV is identified and its spillover mechanisms and transmission pathways are fully characterized. 1. Cui J, Han N, Streicker D, Li G, Tang X, Shi Z, Hu Z, Zhao G, Fontanet A, Guan Y, Wang L, Jones G, Field HE, Daszak P, Zhang S (2007) Evolutionary relationships between bat coronaviruses and their hosts. Emerg Infect Dis 13:1526–1532 2. Fenn TD, Ringe D, Petsko GA (2003) POVScript+: a program for model and data visualization using persistence of vision ray-tracing. J Appl Crystallogr 36:944–947 3. Jones TA, Zou JY, Cowan SW, Kjeldgaard M (1991) Improved methods for building protein models in electron density maps and the location of errors in these models. Acta Crystallogr A 47:110–119 4. Ksiazek TG, Erdman D, Goldsmith CS, Zaki SR, Peret T, Emery S, Tong S, Urbani C, Comer JA, Lim W, Rollin PE, Dowell SF, Ling AE, Humphrey CD, Shieh WJ, Guarner J, Paddock CD, Rota P, Fields B, DeRisi J, Yang JY, Cox N, Hughes JM, LeDuc JW, Bellini WJ, Anderson LJ (2003) A novel coronavirus associated with severe acute respiratory syndrome. N Engl J Med 348:1953–1966 5. Lau SK, Woo PC, Li KS, Huang Y, Tsoi HW, Wong BH, Wong SS, Leung SY, Chan KH, Yuen KY (2005) Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc Natl Acad Sci USA 102:14040–14045 6. Li F, Li W, Farzan M, Harrison SC (2005) Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science 309:1864–1868 7. Li F (2008) Structural analysis of major species barriers between humans and palm civets for severe acute respiratory syndrome coronavirus infections. J Virol 82:6984–6991 8. Li W, Shi Z, Yu M, Ren W, Smith C, Epstein JH, Wang H, Crameri G, Hu Z, Zhang H, Zhang J, McEachern J, Field H, Daszak P, Eaton BT, Zhang S, Wang LF (2005) Bats are natural reservoirs of SARS-like coronaviruses. Science 310:676–679 9. Li W, Zhang C, Sui J, Kuhn JH, Moore MJ, Luo S, Wong SK, Huang IC, Xu K, Vasilieva N, Murakami A, He Y, Marasco WA, Guan Y, Choe H, Farzan M (2005) Receptor and viral determinants of SARS-coronavirus adaptation to human ACE2. EMBO J 24:1634–1643 10. Li W, Wong SK, Li F, Kuhn JH, Huang IC, Choe H, Farzan M (2006) Animal origins of the severe acute respiratory syndrome coronavirus: insight from ACE2-S-protein interactions. J Virol 80:4211–4219 11. Peiris JS, Lai ST, Poon LL, Guan Y, Yam LY (2003) Coronavirus as a possible cause of severe acute respiratory syndrome. Lancet 361:1319–1325 12. Ren W, Li W, Yu M, Hao P, Zhang Y, Zhou P, Zhang S, Zhao G, Zhong Y, Wang S, Wang LF, Shi Z (2006) Full-length genome sequences of two SARS-like coronaviruses in horseshoe bats and genetic variation analysis. J Gen Virol 87:3355–3359 13. Ren W, Qu X, Li W, Han Z, Yu M, Zhou P, Zhang SY, Wang LF, Deng H, Shi Z (2008) Difference in receptor usage between severe acute respiratory syndrome (SARS) coronavirus and SARS-like coronavirus of bat origin. J Virol 82:1899–1907 14. Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596–1599 15. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997) The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25:4876–4882 16. Tu C, Crameri G, Kong X, Chen J, Sun Y, Yu M, Xiang H, Xia X, Liu S, Ren T, Yu Y, Eaton BT, Xuan H, Wang LF (2004) Antibodies to SARS coronavirus in civets. Emerg Infect Dis 10:2244–2248 17. Yu M, Stevens V, Berry JD, Crameri G, McEachern J, Tu C, Shi Z, Liang G, Weingartl H, Cardosa J, Eaton BT, Wang LF (2008) Determination and application of immunodominant regions of SARS coronavirus spike and nucleocapsid proteins recognized by sera from different animal species. J Immunol Methods 331:1–12 18. Yuan J, Hon CC, Li Y, Wang D, Xu G, Zhang H, Zhou P, Poon LL, Lam TT, Leung FC, Shi Z (2010) Intraspecies diversity of SARS-like coronaviruses in Rhinolophus sinicus and its implications for the origin of SARS coronaviruses in humans. J Gen Virol 91:1058–1062 This work was jointly funded by the State Key Program for Basic Research Grants (2005CB523004, 2010CB530100) from the Chinese Ministry of Science, Technology and the Knowledge Innovation Program Key Project administered by the Chinese Academy of Sciences (KSCX1-YW-R-07) to Z.S. and the CSIRO CEO Science Leader Award to L.-F.W. We thank Gary Crameri and Jennifer Barr for help with live virus infection studies. Yuxuan Hou , Cheng Peng , Yan Li , Zhenggang Han & Zhengli Shi Meng Yu & Lin-Fa Wang Fang Li Correspondence to Lin-Fa Wang or Zhengli Shi. Below is the link to the electronic supplementary material. Hou, Y., Peng, C., Yu, M. et al. Angiotensin-converting enzyme 2 (ACE2) proteins of different bat species confer variable susceptibility to SARS-CoV entry. Arch Virol 155, 1563–1569 (2010). https://doi.org/10.1007/s00705-010-0729-6 Received21 April 2010 Accepted12 June 2010 Published22 June 2010 Issue DateOctober 2010 Anyone you share the following link with will be able to read this content: Salt Bridge Severe Acute Respiratory Syndrome ACE2 Gene Pseudotype Virus Severe Acute Respiratory Syndrome Coronavirus *****************************
*****************************
******************************
|























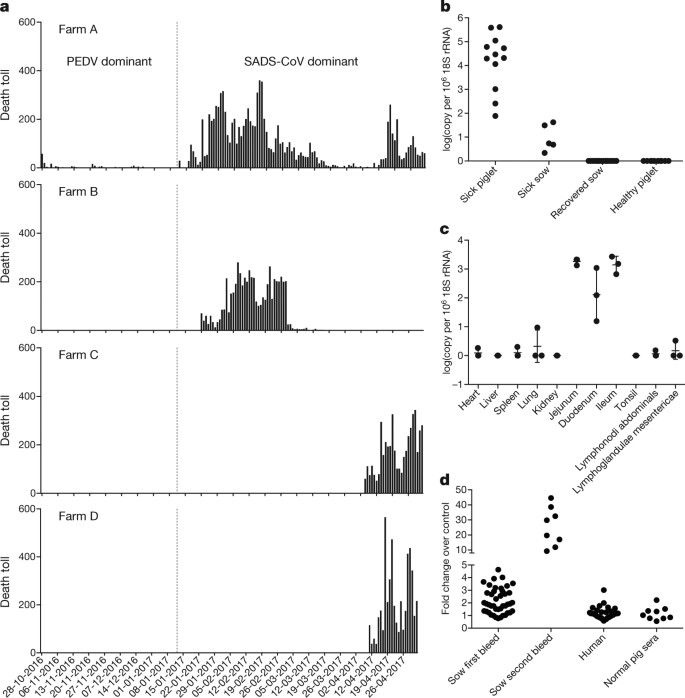
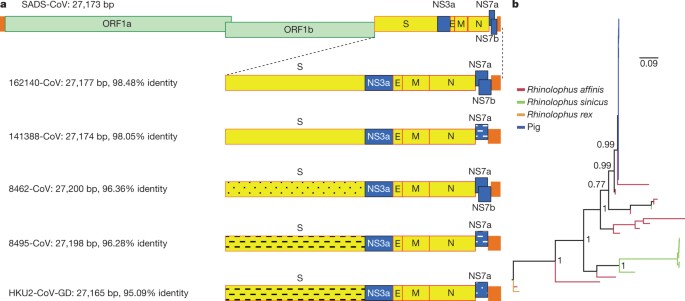
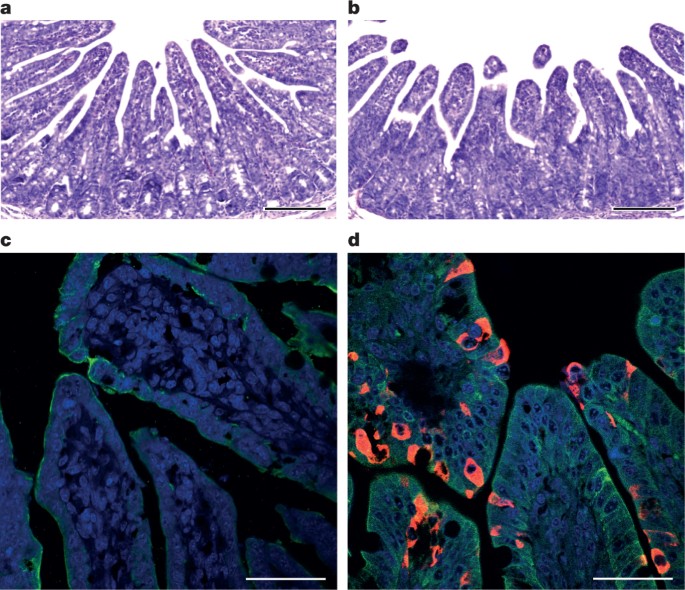












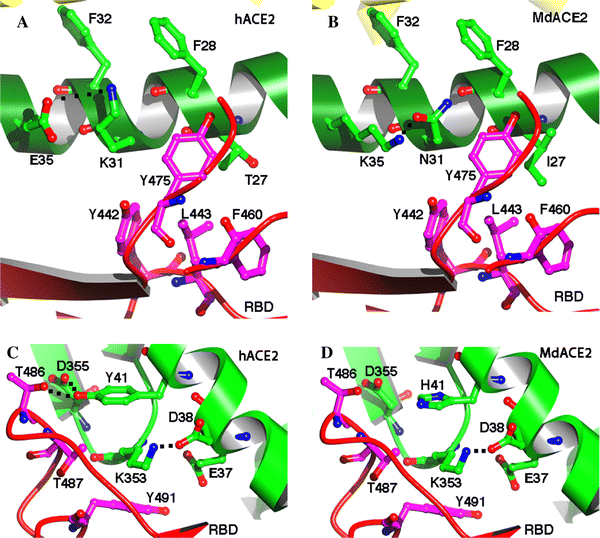

























|
|
|
|
 |
 |
| 實用資訊 | |
|
|
|
|
| 一周點擊熱帖 | 更多>> |
|
|
|
| 一周回復熱帖 |
|
|
|
|
| 歷史上的今天:回復熱帖 |
| 2019: | 吉祥源考 -- 學佛兼學老莊史記資治通鑑 | |
| 2019: | 加稅吧,3月1日期限沒什麼意義了,長期 | |
| 2018: | 攀登乞力馬扎羅山 | |
| 2018: | 躲過兵役的人要閱兵,看看大家的議論 | |
| 2017: | 看沒看央視的新聞。75%的篇幅講兩件事 | |
| 2017: | CNN評論員指出法律站在川普一邊。 | |
| 2016: | 紅樹林,請通告茶壇把A_123給封了!證 | |
| 2016: | 真是好玩:春晚神評 | |
| 2015: | 這次給俺爹做手術,翻他的醫保卡醫療本 | |
| 2015: | 美國高速路上之撞車--智、技、識不如人 | |
