接续:谁设计、制造了新冠病毒(三 上)
上篇文章指出:新冠病毒来自实验室,它集多种冠状病毒、非冠状病毒结构、特性于一身,是广为借鉴、精心设计的跨科属病毒集大成者;新冠病毒出自长期从事功能增益研究的病毒学家、实验室之手。
上篇文章还指出:
1、新冠病毒决定ACE2结合能力的RBD(Receptor Binding Domain,受体结合域)关键氨基酸,与蝙蝠冠状病毒rs3367(WIV1)的RBD关键氨基酸高度关联:新冠病毒5个RBD关键氨基酸中的三个与rs3367(WIV1)的对应RBD关键氨基酸相同(第二、第四、第五RBD关键氨基酸),此外,二者还有一对RBD关键氨基酸理化属性高度接近(第三RBD关键氨基酸)。

2、新冠RBD与rs3367(WIV1)RBD空间结构非常相似(可能高度同构),二者RBD-ACE2结合模型的空间构象也非常相似(可能高度同构)。
3、新冠、rs3367(WIV1)都有非常强大的人体细胞进入、感染能力,它们的刺突蛋白都非常适合结合hACE2(human ACE2)。
4、新冠、rs3367(WIV1)都有非常强大的跨物种感染能力和广泛的宿主范围。
5、新冠、rs3367(WIV1)都有泛器官、泛组织感染能力。
6、新冠与rs3367(WIV1)的上述关联、“巧合”不是自然演化产生的,它们是人为设计的结果,有人借鉴rs3367(WIV1)的RBD关键氨基酸,通过氨基酸复用、替代,设计出了结构、功能高度相似、高度等价的新冠病毒RBD关键氨基酸。
借鉴rs3367(WIV1),设计出新冠病毒RBD关键氨基酸的,是哪个病毒学家,哪个实验室?
本系列首篇文章
谁设计、制造了新冠病毒(一) (即“新冠病毒与拉尔夫·巴里克的不解之缘(一)”)
及另一篇文章
谁设计、制造了新冠病毒(二 下)
分别指出,新冠病毒逆转录自身RNA为cDNA(RNA的互补DNA),并将其整合到人体被感染细胞DNA中的特性,及新冠病毒S1/S2处的“RRAR”弗林(furin)酶切位点结构,这两项人为设计的重大嫌疑人,都是北卡罗来纳大学教堂山分校微生物学和免疫学院流行病学系教授、冠状病毒顶级研究权威、冠状病毒“合成之父”,冠状病毒功能增益改造狂人,现美国国家科学院院士拉尔夫·S·巴里克(Ralph S. Baric)。
新冠病毒逆转录RNA-DNA整合特性,新冠病毒furin酶切位点结构,与拉尔夫·S·巴里克论文内容、发明内容、项目内容的关联,没准都是偶然的巧合,新冠病毒的RBD关键氨基酸与Ralph S. Baric应该不会再有密切关联了。
有个著名病毒学家,他及他领导的科学团队:
1、对冠状病毒刺突蛋白、RBD、RBD关键氨基酸,对它们与不同物种ACE2,特别是与hACE2的作用机制研究得极为精深、透彻;
2、有着丰富的冠状病毒功能增益改造经验(10多年),丰富的冠状病毒合成、改造经验(约20年,2000年就发明了基于基因序列无痕迹合成、改造冠状病毒的反向遗传平台);
3、曾以不能感染人类的宿主单一的动物冠状病毒(果子狸冠状病毒)刺突蛋白为原材料,通过RBD关键氨基酸的替代改造、培育变异,改造、培育出了有人类感染、致病能力,可跨物种感染、传播的冠状病毒(2007年);
4、多次研究WIV1(rs3367),对其结构、特性,包括RBD关键氨基酸,hACE2结合能力,人体细胞感染能力了如指掌;
5、曾以WIV1的刺突蛋白为功能增益改造的原材料,制造出了极易结合hACE2,人体细胞感染能力极强的高危病毒(WIV1-MA15,2016年)。
这个对rs3367(WIV1),对RBD关键氨基酸都做过深入研究的人是谁呢?拉尔夫·S·巴里克,拉尔夫·S·巴里克是唯一一个同时符合五项描述的(不计括号内的补充描述)病毒学家。
与拉尔夫·巴里克存在重大关联的新冠病毒人为设计特征不只上述三组:
逆转录RNA-DNA整合特性(逆转录是拉尔夫·S·巴里克发明的反向遗传平台的最基本技术,也是使用反向遗传平台合成冠状病毒的必要步骤)、
S1/S2处的furin酶切位点(在2018年初美国国防部高级研究计划局启动,彼得·达萨克-生态健康联盟代为组织的defuse项目中,拉尔夫·S·巴里克负责刺突蛋白的反向遗传工程改造,其中一项改造内容是在蝙蝠冠状病毒刺突蛋白中插入furin酶切位点)、
RBD关键氨基酸,稍后的“论文A-2”部分还将指出第四组极为“巧合”的关联:2016年3月拉尔夫·巴里克团队PNAS(美国国家科学院院刊)论文中展望、预测的刺突蛋白RBD外区域的三项改造成效,在三年后出现的新冠病毒中全都神奇地实现了!
新冠病毒与拉尔夫·巴里克的论文内容、发明内容、项目内容间还存在其它结构、特性的可疑关联、“巧合”,本文不继续展开了。
我认为,拉尔夫·巴里克就是新冠病毒的核心设计者,拉尔夫·巴里克(团队)不是新冠病毒核心设计者的可能性微乎其微。
(补充说明一下,新冠病毒的结构、特性可能并非全部出自拉尔夫·巴里克一人(一支团队)之手,新冠病毒可能是跨实验室设计、改造的产物。拉尔夫·巴里克曾将新冠病毒原型共享给其它实验室,该实验室可能对新冠病毒原型做过二次改造。)
拉尔夫·巴里克对功能增益改造(gain-of-function)研究狂热而执着。奥巴马禁令(即“流感、SARS、MERS相关的危险病原体功能增益研究暂停令”,奥巴马政府2014年10月17日颁布)颁布后,拉尔夫·巴里克大为不满,他给NIH(National Institute of Health,美国国立卫生研究院,当时院长为弗朗西斯·柯林斯(Francis S. Collins))写了一封长信,称暂停令将严重影响冠状病毒研究,如果未来再爆发疫情,科学家将不能及时应变,迅速控制疫情。他要求NIH批准他继续开展两项SARS相关的功能增益研究并得到了NIH的特批(奥巴马禁令中有特批的条款)。完成了这两项研究后,拉尔夫·巴里克团队先后发表了2015年11月9日的Nature Medicine论文和2016年3月14日的PNAS论文,这两篇论文就是稍后将介绍的论文A-1和论文A-2。
功能增益研究打着模拟、预测冠状病毒自然突变,防患于未然的旗号,听上去冠冕堂皇;拉尔夫·巴里克(团队)的众多论文其实是在探索:
如何改变冠状病毒的感染嗜性和宿主对象,如何将不能感染人类的动物来源的冠状病毒改造为可感染人类的冠状病毒;
如何将不会使人发病的动物来源的冠状病毒(少数蝙蝠冠状病毒有人体细胞进入、感染能力,但已知的所有蝙蝠冠状病毒都不会使人体产生临床疾病症状)改造为有人类致病能力的冠状病毒;
如何改变、扩大冠状病毒的宿主范围,如何将宿主单一的动物来源的冠状病毒改造为可跨物种感染、传播的冠状病毒;
一句话,拉尔夫·巴里克在长期研究、探索改变、增强冠状病毒感染、致病、传播能力的技术。
功能增益改造研究不能防患于未然,它带来的不是人类的安全,而是人为灾难。功能增益研究可以随心所欲地改造病毒基因,轻易突破、跨越大自然设置的病毒感染物种隔离壁垒,轻松揭掉潘朵拉灾难魔盒上的封印。与病毒感染、致病、传播能力有关的保守基因的关键突变,在自然条件下极难发生,但是,自然条件下数十万年、数百万年、数千万年。。。都不会发生的基因改变,在拉尔夫·巴里克这样的功能增益改造高手的实验室中,几天、甚至若干小时就可以制造出来了。
即使面对着悬崖,即使声音无比微弱,即使难得回响,我也要尽力喊出来。我们只有被欺瞒、被愚弄的命吗?
下面介绍拉尔夫·巴里克研究rs3367(WIV1)、RBD 关键氨基酸的一些论文实例。
Ralph S. Baric对WIV1(rs3367)的研究
Ralph S. Baric对rs3367(WIV1)非常关注,2015年到2018年四年间,其团队先后发表了四篇与WIV1有关的论文,在其中两篇论文中,WIV1是病毒主角,论文的专门研究对象。
论文A-1
2015年11月9日,Ralph S. Baric领导的团队在《自然医学》(Nature Medicine)杂志发表了一篇现在广为人知的冠状病毒嵌合改造论文:A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence(一个类似SARS的蝙蝠冠状病毒群显示了产生人类流行疫情的潜力)
https://www.nature.com/articles/nm.3985
论文将蝙蝠冠状病毒SHC014(也叫rsSHC014)的刺突蛋白与SARS-CoV MA15的骨架嵌合,制造出了一种高度危险的实验室病毒SHC014-MA15。嵌合病毒SHC014-MA15具有跨物种感染能力,能强烈感染Vero E6细胞(非洲绿猴肾细胞系细胞)和HAE细胞(human airway epithelium cells,人类气道上皮细胞),它在这两种细胞中的复制滴度(大致相当于复制浓度)与SARS流行病毒株相当;SHC014-MA15毒性很强,能使实验小鼠体重大幅下降并死亡;SARS单克隆抗体和SARS疫苗都不能有效治疗SHC014-MA15的感染。
SARS-CoV MA15是Ralph S. Baric等人在实验室小鼠体内反复传代培育出的SARS病毒小鼠适应性变异体,对小鼠有极强的毒性和100%的致死力。原生的人类流行SARS病毒对人类致病能力很强,但对小鼠致病能力很弱。
WIV1在论文中是病毒主角SHC014的对照病毒(SHC014、rs3367、WIV1都发现于云南同一山洞)。论文指出:
在决定ACE2结合能力、细胞进入能力、宿主范围的5个RBD关键氨基酸(残基)中,WIV1虽然有三个关键氨基酸与SARS不同(二者第3、5关键氨基酸相同,第1、2、4关键氨基酸不同,见下图),但它的刺突蛋白仍能结合hACE2,WIV1和SARS一样,也具有人体细胞进入(感染)能力。
注:WIV1具有人体细胞进入、感染能力,是武汉所2013年10月Nature论文(https://www.nature.com/articles/nature12711)的研究结论,论文A-1引用了这一结论。2013年武汉所论文的核心实验就是证明WIV1具有人体细胞进入、感染能力。
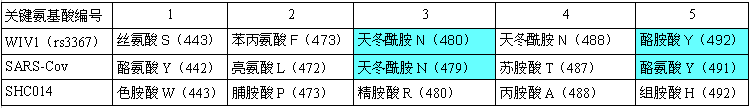
SARS、WIV1、SHC014 RBD关键氨基酸对照表
论文随后把目光转向SHC014,论文指出,相比WIV1,SHC014与SARS的RBD关键氨基酸差异更大,SHC014的RBD关键氨基酸无一与SARS 相同,但是,仍不能断言其刺突蛋白没有hACE2结合能力。论文用SHC014的刺突蛋白构建嵌合病毒SHC014-MA15的首要目的,就是用SHC014-MA15来实验确定SHC014刺突蛋白的hACE2结合能力。
论文A-1经常被人张冠李戴,说成是石正丽团队发表的论文。关于论文主创、作者,及石正丽团队成员有否参加相关研究,下文“错误一”章节中有细致的说明:
改造病毒问题兼批驳王文清新冠溯源文章
论文A-2
2016年3月14日,Ralph
S. Baric领导的团队在PNAS(美国国家科学院院刊)上发表了另一篇重要论文:SARS-like WIV1-CoV poised for
human emergence(类SARS冠状病毒WIV1-CoV对人类有潜在威胁)
https://www.pnas.org/content/113/11/3048
这是一篇专门研究WIV1的论文,上篇文章也提到了该论文。
论文用反向遗传平台合成了WIV1的克隆,称之为WIV1-CoV;同时用WIV1的刺突蛋白与SARS-CoV MA15的骨架合成了嵌合病毒WIV1-MA15。
体外细胞实验证明,和论文A-1中的SHC014-MA15一样,WIV1-MA15也能强烈感染灵长类动物的Vero E6细胞(非洲绿猴肾细胞)和人气道上皮细胞(HAE细胞),它在这两种细胞中的复制滴度也与SARS-CoV流行病毒株相当。
小鼠体内实验证明,WIV1-MA15、WIV-CoV能感染实验小鼠,在小鼠气道和肺中有效复制,但它们在小鼠肺中的复制滴度比之小鼠适应性变异体SARS-CoV MA15低10~100倍;它们对小鼠的致病力都比较弱,只能使年轻小鼠(10周龄)产生较轻症状,不会使其体重显著下降(超过10%),更不会使其致死。
但是,当感染对象由普通实验小鼠换成表达hACE2(human ACE2)的转基因实验小鼠(其肺、脑、肝、肾和胃肠道等部位的mACE2被转基因为hACE2)后,WIV1-MA15和WIV-CoV的感染、复制、致病能力大大增强。相比普通小鼠,WIV1-MA15、WIV1-CoV在转基因小鼠肺中的复制滴度提高了100倍;部分10-20周龄的被感染转基因小鼠体重减轻超过10%;两种病毒可使转基因小鼠致死,它们在转基因小鼠大脑中的复制更强劲(与肺中相比),使部分转基因小鼠患上了致命的脑炎。
转基因小鼠体内感染实验表明,hACE2是WIV1刺突蛋白的极佳结合对象;如果要构造具有强大人体细胞感染能力的冠状病毒,那么WIV1的刺突蛋白、RBD、RBD关键氨基酸是极佳的原材料、极佳的参照物。
论文Discussion部分指出:冠状病毒刺突蛋白RBD外区域的变化,如S2亚基的某些变化,S1亚基RBD外区域的某些变化(RBD是S1亚基的一部分),有可能增强刺突蛋白与宿主蛋白酶(ACE2受体就是一种宿主蛋白酶)作用的靶向性(可理解为针对性、目的性),增进刺突蛋白的切割(即酶切)特性,增进刺突蛋白的扩展性。这实际上指出了对刺突蛋白RBD外区域进行改造可能取得的三项收效,令人难以置信的是,论文展望、预测的这三项RBD外区域改造成效,在3年又8个月后出现的新冠病毒中,全都神奇地实现了!详情请参阅:
科学疯子设计的病毒集大成者(二)
Ralph S. Baric团队是唯一一支将WIV1(rs3367)用于功能增益改造的科学团队,是唯一一支以WIV1(rs3367)为原材料改造出人类致病病毒的科学团队。
论文A-1、A-2,即Ralph S. Baric团队2015年11月以SHC014为病毒主角的nature medicine论文,2016年3月以WIV1为病毒主角的PNAS论文都发表于奥巴马禁令生效期间,两篇论文对应的两项SARS相关的功能增益研究(都以SARS小鼠适应性变异体SARS-CoV MA15为嵌合病毒的骨架)都在NIH的特批下得以继续实施、完成并发表论文。2017年12月19日,川普政府撤销了奥巴马暂停令,全面重启了美国危险病原体功能增益研究,重新为从事该类研究的科学家、实验室提供联邦资金资助。美国举国性质的危险病原体功能增益研究重启两年后,2019年11月、12月,新冠病毒在武汉出现,新冠全球疫情爆发。
论文A-3
2017年6月28日,Ralph
S. Baric团队(北卡罗来纳大学教堂山分校流行病学系)、范德堡大学医学中心、波兰Jagiellonian
University、吉利德科技等在《ScienceTranslational
Medicine》杂志联合发表了如下论文:Broad-spectrum antiviral GS-5734 inhibits both
epidemic and zoonotic coronaviruses(广谱抗病毒药物 GS-5734可抑制流行性人畜共患冠状病毒)
https://stm.sciencemag.org/content/9/396/eaal3653
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5567817/
GS-5734是瑞德西韦(Remdesivir)的研发编号。
论文指出:目前正在临床开发的用于治疗埃博拉病毒的GS-5734对多种冠状病毒,包括SARS-CoV和MERS-CoV都有明显的抑制疗效,GS-5734的预防性早期治疗显著降低了肺病毒载量,改善了临床症状和呼吸功能。
为验证GS-5734即瑞德西韦的广谱抗(冠状)病毒能力,Ralph S. Baric等人使用反向遗传平台,基于病毒的基因序列合成了7种冠状病毒,WIV1是其中之一,其它六种病毒是:
SARS-CoV(SARS病毒)、
MERS-CoV(中东呼吸综合症病毒)、
HCoV-NL63(人类冠状病毒NL63)、
HKU3(蝙蝠冠状病毒HKU3)、
HKU5(伏翼蝠冠状病毒HKU5)、
SHC014(蝙蝠冠状病毒rsSHC014,论文A-1的病毒主角)。
(伏翼蝙蝠是一种拇指大小、重约4克的蝙蝠,发现于2006年,可能是香港独有的物种。)
实验证明,随着GS-5734使用剂量(浓度)的增加,HAE细胞培养物中MERS、SARS、WIV1等7种病毒的RNA滴度均显著下降;
(HAE细胞,即human airway epithelium cells,人气道上皮细胞)
另一组实验证明,随着GS-5734使用剂量(浓度)的增加,7种病毒的两种辅助蛋白 ORF1、ORFN的RT-PCR检测值都显著下降。
论文还分两组比较了WIV1等5种病毒与SARS,以及5种病毒与MERS在nsp12蛋白(一种辅助蛋白)部分、及spike蛋白(刺突蛋白)部分的A.A. identity(Amino Acid identity,氨基酸序列一致度或相似度),比较结果表明:不同冠状病毒的spike蛋白部分差异较大,但它们的nsp12蛋白部分则高度保守(一致程度很高,差异度很低)。因而,(高度一致的)nsp12蛋白是广谱抗冠状病毒药物的适宜靶点(治疗作用点)(nsp12蛋白应该就是GS-5734的治疗靶点)。
论文A-4
2018年12月19日,Ralph S. Baric团队与国家过敏和传染病研究所(所长即安东尼·福奇Anthony Fauci)的两个下属机构(病毒学实验室、落基山兽医分会)在著名国际开源出版期刊MDPI(Multidisciplinary Digital Publishing Institute)上联合发表了如下论文:SARS-Like Coronavirus WIV1-CoV Does Not Replicate in Egyptian Fruit Bats(类SARS冠状病毒WIV1不会在埃及果蝠中复制)
https://www.mdpi.com/1999-4915/10/12/727/htm
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6316779/
这是Ralph S. Baric另一篇专门研究WIV1的论文。
论文通过果蝠体内实验指出:WIV1病毒能有效利用埃及果福的ACE2受体进入果蝠细胞,它能感染果蝠的鼻甲、气管、肺、肾、胃、肠等众多器官、组织,使这些器官、组织产生明显的不同程度、不同细节的免疫反应。
注:埃及果蝠(Egyptian fruit bat,Rousettus aegyptiacus)是马尔堡病毒的天然宿主,也是所有丝状病毒的唯一已知宿主。
上篇文章曾指出,WIV1病毒具有泛器官、泛组织感染能力,依据就是这篇论文。
WIV1虽然可感染果蝠多处器官、组织,并引发免疫反应,但它的毒性很低。实验证明,WIV1不会对果蝠造成强烈感染,不会引发可观察的(外部)临床症状,12只实验果蝠都没有表现出明显的临床疾病迹象(如呼吸窘迫、厌食或嗜睡),未检测到果蝠的体重减轻或体温变化。论文判断:WIV1可能在果蝠体内只发生了一些低水平的病毒复制。
这表明,WIV1虽然具有泛器官、泛组织感染能力,但它(对果蝠)的致病能力非常弱。(对果蝠)它没有表现出新冠病毒那样的泛器官、泛组织损伤、破坏能力(它更不具备新冠病毒所拥有的完备、周全的宿主免疫对抗能力,如免疫屏蔽、免疫逃避,免疫干扰、免疫抑制,免疫破坏等)。
研究者在多个感染后时段对实验果福分批实施安乐死,并对果福的超过20个器官、组织部位进行了病毒学和组织病理学分析:鼻甲、喉、咽、气管、肺、脑、眼、结膜、心、肝、脾、肾、膀胱、生殖器官、胃、近端和远端肠道、颈部淋巴结、肾上腺、皮肤和骨骼肌。
这表明,Ralph S. Baric团队清楚地认识到了WIV1病毒的泛器官、泛组织感染能力。
感染后第三天的检测、分析结果是:除在果蝠咽部和鼻甲骨检测到WIV1病毒RNA外,在其它器官、组织中均未再检测到WIV1病毒RNA。
也就是说,WIV1病毒进入、感染了众多果蝠器官、组织,引发了这些器官、组织的(内部)免疫反应,但几天之后,它又从绝大多数器官、组织中消失了。论文认为,WIV1消失的原因是它在果福器官、组织中缺乏持续、有效的复制、繁衍。
论文还做了两组转基因BHK细胞(Baby Hamster Syrian Kidney cells,叙利亚幼仓鼠肾细胞)体外感染实验,实验结果表明,WIV1能有效感染表达hACE2的转基因BHK细胞,和表达果蝠ACE2 的转基因BHK细胞;与果福体内感染实验不同的是,WIV1在这两种转基因BHK细胞中都能健壮地持续复制。
Ralph S. Baric不只对WIV1(rs3367)做过反复研究,他的几乎每一篇论文(我读过的)都要研究冠状病毒的刺突蛋白,刺突蛋白的RBD;对RBD关键氨基酸,Ralph S. Baric早在2007年就做过极为精深的研究。下文是4个RBD关键氨基酸论文实例:
拉尔夫·巴里克对RBD关键氨基酸的研究
(未完待续)






