他们扮瞎、装聋、作哑,不调查、不讨论、不声张、不宣传,不回应、不澄清,不等于它们不存在。
笔者相信,有良知,有正义感、责任感,有铁肩道义的媒体人不会回避、冷处理、冷漠对待理性客观,逻辑严谨,证据坚实、充分,内容翔实、深刻,不可多得的溯源作品,不会对新冠病毒诸多隐性、长期的重大危害闭目塞听、无动于衷。笔者相信,有良知、有责任感的媒体人会热情鼓励对新冠来源真相的探查、挖掘、揭示,会高度关注,积极展示、传播溯源的重大发现、重大证据、重大线索、重大疑点,会大力支持溯源研究、讨论、争鸣和观点对撞。
冠状病毒拥有最大的RNA基因组,可提供充裕的基因改造空间,便于在同一病毒中引入、模仿、拼接、叠加、整合、集成种种来源的跨物种基因,在同一病毒中实现、汇集各种各样的感染、致病、免疫对抗(免疫破坏、免疫逃避和免疫屏蔽、免疫干扰和免疫抑制)机制。冠状病毒是理想的基因工程、基因改造对象,是制造超级病原体的最佳底版。
SARS-CoV-2继承、借鉴、整合、汇集了多种冠状病毒的基因、蛋白结构;
SARS-CoV-2引入、模仿、汇集了大量各科属非冠状病毒的基因、蛋白、抗原结构;
SARS-CoV-2引入、模仿了众多非病毒微生物(致病细菌和微生物寄生虫)的基因、蛋白、抗原结构;
SARS-CoV-2含有多个与人类完全相同或高度相似,并在感染、致病中发挥重要作用的基因、蛋白结构,SARS-CoV-2还具有其它一些模仿人类蛋白、多肽结构及特定功能的基因、蛋白结构;
SARS-CoV-2含有某些低等动物特有的基因、蛋白结构;
SARS-CoV-2含有植物特有的基因、蛋白/多肽结构;
SARS-CoV-2含有与细菌(分泌)毒素、动物(分泌)毒素高度相似或功能相同的基因序列/氨基酸序列;
SARS-CoV-2还含有与医疗药剂高度相似的基因序列/氨基酸序列,以及与生物医学专利完全匹配的基因序列。
SARS-CoV-2是广为参照、借鉴(包括不加改动地抄袭、复用重要序列)、广为移植、模仿,广为优化、整合的基因工程恢弘巨制,是身披冠状病毒躯壳的跨物种基因、跨生物界基因、超生物基因聚合体,是病原体转基因改造集大成者。
I SARS-CoV-2对冠状病毒基因的继承、借鉴、模仿、优化、集成
CoVZC45/CoVZXC21
蝙蝠冠状病毒CoVZC45或CoVZXC21为SARS-CoV-2提供了冠状病毒躯壳、主要病毒骨架和改造底板。
CoVZC45、CoVZXC21从中国浙江舟山的小菊头蝠样本中分离获得,样本收集时间分别为2017年2月、2015年7月,它们的全基因组序列上传并实现国际共享的时间都是2018年1月。这两种蝙蝠冠状病毒高度相似,二者全基因组序列一致性约为97.48%。
SARS-CoV-2与CoVZC45/CoVZXC21多种蛋白高度相似。

CoVZC45、CoVZXC21、SARS-CoV与SARS-CoV-2的基因、蛋白一致性
注:图中的SARS-CoV GZ02是SARS疫情(2002年11月-2003年9月)早期于广州收集、鉴定的SARS流行病毒株。
由图可见,SARS-CoV-2与CoVZC45 E蛋白(Envelope protein,包膜蛋白)aa序列(amino acid sequence,氨基酸序列)一致性(identity,同一性/一致度/相似度)为100%,M蛋白(Membrane protein,膜蛋白)一致性为98.6%,二者N蛋白(Nucleocapsid protein,核衣壳蛋白)、ORF1ab(open reading frame 1ab,开放阅读框1ab)多蛋白基因,以及ORF3、ORF7、ORF10、ORF14等辅助蛋白的一致度也都大于90%。二者S蛋白(Spike protein,刺突蛋白)、ORF9b(旧称ORF13)蛋白的一致度较低,分别为80.2%、73.2%,与其它蛋白的一致度存在较大反差。
SARS-CoV-2与CoVZC45全基因组序列(核苷酸序列)一致度约为87.6%,与CoVZXC21全基因组序列一致度约为87.5%。
SARS-CoV-2、CoVZC45多种蛋白一致度大于90%,但S蛋白的一致度仅为80.2%,这意味着SARS-CoV-2的S蛋白经历了较多改造,引入了更多非CoVZC45基因。两种病毒S蛋白两个亚基的一致度更不均衡,二者S蛋白S2亚基(膜融合亚基,spike的后半部分)的一致性高达95%,而S1亚基(受体结合亚基,spike的前半部分)的一致性却低至69%。近亲病毒间同一蛋白组件相似度如此不协调,反差如此巨大,这对自然演化来说极为反常的,这表明,SARS-CoV-2 S1亚基经历的基因编辑、改造远多于S2亚基。
SARS-CoV
SARS-CoV是SARS-CoV-2 的另一个重要参照模板,它与CoVZC45/CoVZXC21共同作为SARS-CoV-2的基本病毒骨架、基本改造底板。
由前一小节中的配图可见,SARS-CoV与SARS-CoV-2全基因组序列、各蛋白基因序列(核苷酸序列)、各蛋白aa序列的一致性皆低于CoVZC45/CoVZXC21(ORF1b部分略高于CoVZXC21);然而,SARS-CoV-2 的感染、致病能力、特性却与SARS-CoV,而非CoVZC45/CoVZXC21相似,CoVZC45、CoVZXC21虽然“宏观上”与SARS-CoV-2更相像,但二者都没有人类感染、致病能力。
蝙蝠因携带多种冠状病毒而成为人祸的最佳背锅对象。有必要指出,已知的蝙蝠冠状病毒无一具有人类致病能力。少数蝙蝠冠状病毒虽然具有ACE2结合能力和人体细胞进入能力,但它们在人体内不能有效复制,不能持续存在;截止2020年,已知的人类致病冠状病毒只有7种,它们与蝙蝠冠状病毒可能存在进化渊源,但它们自身都不是蝙蝠冠状病毒;已知的蝙蝠冠状病毒无一可使人体产生临床疾病症状。真正危险、邪恶的不是蝙蝠或蝙蝠冠状病毒,而是人,对蝙蝠冠状病毒实施G-o-F(Gain-of-Function)改造的人,介入、支持改造研究的人,对真相心知肚明但声称基因改造病毒是蝙蝠冠状病毒自然进化产生的人。
SARS-CoV-2、SARS-CoV全基因组序列一致性约为79%,Spike蛋白aa序列一致性约为76.47%,Spike蛋白S1亚基aa序列一致性约为64.85%,S1亚基RBD(receptor-binding domain,受体结合域)aa序列一致性约为73%–76%,二者RBD决定ACE2结合能力的五个关键氨基酸仅有一个相同。可见,虽然互为“近亲”,但SARS-CoV-2与SARS-CoV差异很大,通过自然演化来消除两者间的差异,也许需要上百万年、上千万年,然而,从2002年SARS-CoV突然出现之后又突然消失,到2019年SARS-CoV-2出现,只用了17年。没有可作为它们共同祖先的病毒,没有过渡衔接的病毒。
尽管差异很大,尽管进化距离非常遥远,但是,SARS-CoV-2许多关键/重要的感染、致病结构,感染、致病能力继承或优化自SARS-CoV。
1、SARS-CoV-2继承、提升了SARS-CoV的ACE2结合能力。
SARS-CoV-2、SARS-CoV的Spike蛋白都具有很强的ACE2结合能力,并且,二者Spike蛋白S1亚基与ACE2的结合复合体非常相似。相比之下,CoVZC45、CoVZXC21都不具备有效的ACE2结合能力和人体细胞进入能力。
SARS-CoV-2 的ACE2结合能力比SARS-CoV更强大,德克萨斯大学奥斯汀分校的一项研究指出,SARS-CoV-2 Spike的ACE2结合亲和力比SARS-CoV Spike高10-20倍(也有研究认为,SARS-CoV-2 RBD的ACE2结合亲和力高于SARS-CoV RBD,但Spike整体/full-length spike的ACE2结合亲和力与SARS-CoV-2 Spike相当或略低)。下一篇文章将说明,SARS-CoV-2决定ACE2结合能力的五个RBD关键氨基酸借鉴自蝙蝠冠状病毒WIV1(rs3367)。
2、SARS-CoV-2继承、优化了SARS-CoV与HIV-1高度相似的膜融合蛋白和膜融合介导机制。
HIV-1(human immunodeficiency virus-1,人类免疫缺陷病毒-1/ 艾滋病病毒)是逆转录科慢病毒属病毒,它是SARS-CoV-2最重要的非冠状病毒参照、借鉴、模仿对象,SARS-CoV-2、HIV-1之间存在着惊人的众多相似和关联,其中一部分相似、关联在SARS-CoV-1(SARS-CoV)、HIV-1之间其实已经存在(但几乎不被提及)。
SARS-CoV-1 的膜融合蛋白-S2蛋白(即Spike蛋白的S2亚基)与HIV-1的膜融合蛋白-gp41蛋白之间不存在序列同源性(sequence homology,指显著的序列相似性),然而,gp41蛋白的所有重要元件在S2蛋白中都存在,而且,两种蛋白构造上高度相似。

HIV-1 gp41蛋白(a)、SARS-CoV-1 S2蛋白(b)构造对比图
由上图可见:
1)HIV-1 gp41蛋白、SARS-CoV-1 S2蛋白都含有两个七肽重复区域(heptad repeat,HR)N-HR、C-HR ,S2蛋白的N-HR比gp41蛋白的N-HR更长。两种蛋白的N-HR(N-terminal heptad repeat,N 端七肽重复区域)在图中显示为红棕色,C-HR(C-terminal heptad repeat,C 端七肽重复区域)显示为绿色。
注:N 端,NH2端,氨基端;C端,COOH端,羧基端。
2)在两个七肽重复区域(HR)之间的(半)环上,HIV-1 gp41蛋白、SARS-CoV-1 S2蛋白都具有限制环的发夹构象(hairpin conformation)的半胱氨酸残基(Cysteine residues,C)和脯氨酸残基(Proline residues,P)。gp41蛋白环结构上有2个半胱氨酸残基(C)、1个脯氨酸残基(P);S2蛋白两个HR间的环结构更长,含有4个半胱氨酸残基(C)、9个脯氨酸残基(P)。环结构上的半胱氨酸残基(C)在图中以紫色短线表示,脯氨酸残基(P)在图中未显示。
3)在C-HR(绿色)和跨膜区域/片断(transmembrane segment,橙色)之间,gp41蛋白、S2蛋白都含有一段富含芳香族残基的区域(aromatic residue-rich regions),这一区域在图中显示为蓝色。
在两种蛋白的富含芳香族残基区域,蛋白中通常相对稀有的芳香族残基都约占区域内氨基酸残基总数的一半,如下图所示:

HIV-1 gp41蛋白、SARS-CoV-1 S2蛋白富含芳香族残基区域的aa序列
图中蓝色氨基酸残基W、F、Y为芳香族残基。
W:Tryptophan,Trp,色氨酸;
F:Phenylalanine,Phe,苯丙氨酸;
Y:Tyrosine,Tyr,酪氨酸。
芳香族残基即芳香族氨基酸残基,芳香族氨基酸(Aromatic amino acids)指含有芳香环的氨基酸,包括苯丙氨酸、色氨酸、酪氨酸、组氨酸四种氨基酸。
在前面gp41蛋白、S2蛋白构造对比图中,摆放在gp41蛋白N-HR、C-HR之间的黄色棒体不是gp41蛋白的一部分,它表示抗艾滋病药物Enfuvirtide(恩夫韦肽)。Enfuvirtide,也叫T-20,商品名Fuzeon,1996年开始研发,1999年完成研发,是参照HIV-1 gp41蛋白C 端七肽重复区域(C-HR)aa序列设计的可结合N-端七肽重复区域(N-HR)并有效抑制HIV-1膜融合活性的合成肽(N-HR、C-HR需要相互作用结合形成六螺旋束来介导膜融合,Enfuvirtide通过结合N-HR来竞争、干扰C-HR与N-HR的结合,阻碍六螺旋束的形成)。Enfuvirtide同样可结合SARS-CoV-1 S2蛋白,并被用作SARS-CoV-1 融合抑制剂;它还被证明可强力结合SARS-CoV-2 的S2蛋白,并被建议作为强效SARS-CoV-2融合抑制剂用于COVID-19临床治疗。Enfuvirtide在SARS-CoV-1、SARS-CoV-2 S2蛋白中的作用、结合区域应该也是N-HR(HR1)。
Enfuvirtide的问世可能标识着科学界已经深入认识了HIV-1的膜融合机制;Enfuvirtide研发成功的同一年,Ralph S. Baric等人发明了基于基因序列合成冠状病毒的反向遗传系统,次年(2000)发表了论文;又两年之后的2002年,SARS-CoV出现。
SARS-CoV-1 S2蛋白、HIV-1 gp41蛋白还具有其它相似性。
虽然不存在显著序列相似性(同源性),但两种蛋白两个七肽重复区域N-HR、C-HR的aa序列(amino acid sequence,氨基酸序列)具有相似的螺旋轮结构(helical wheel structure)。螺旋轮结构/螺旋轮图是螺旋轮分析(helical wheel analysis)的结果图,螺旋轮分析是以螺旋轮方式(也是一种密码编码方法)排列蛋白/多肽aa序列中的氨基酸(残基),以表现、分析蛋白/多肽中的α-螺旋。

HIV-1 gp41蛋白N-HR、C-HR aa序列的螺旋轮图

SARS-CoV-1 S2蛋白N-HR、C-HR aa序列的螺旋轮图
此外,SARS-CoV-1 S2蛋白9个连续α螺旋(nine successive alpha-helices)中的2个(对应aa序列片断P879-N942)还与HIV-1 gp41蛋白中的2个α螺旋高度吻合(S2 and gp41 share the same two α helices)。

SARS-CoV S2蛋白(白色)3-D模型、HIV-1 gp41蛋白局部3-D模型叠加图
由图可见,gp41蛋白黄色和蓝色的两个α螺旋与S2蛋白中的两个白色α螺旋高度吻合、重叠。
gp41蛋白是HIV-1 gp160蛋白的两个亚基之一,gp160蛋白是HIV-1的Env蛋白(Envelop protein,包膜蛋白),它在功能上对应冠状病毒的Spike蛋白。gp160蛋白的另一个亚基是gp120蛋白,gp120是受体结合亚基,gp41是膜融合亚基,它们在功能上分别对应冠状病毒Spike蛋白的受体结合亚基-S1亚基、膜融合亚基-S2亚基。
SARS-CoV-1 S2蛋白与HIV-1 gp41蛋白的高度构造相似性可能缘于它们有共同的病毒远祖,或者,可能是自然演化的巧合,SARS-CoV-1 S2蛋白也可能是HIV-1 gp41蛋白的仿制品。
逆转录病毒gp41蛋白、冠状病毒S2蛋白都属于I类融合蛋白(Class I Fusion Proteins),I型病毒融合蛋白具有一些共同特征,如都包含两个七肽重复区域等等;但是,S2蛋白与HIV-1 gp41蛋白的高度构造相似性(至少其中一部分)应该为SARS-CoV-1(、SARS-CoV-2)所专有,两篇指出SARS-CoV-1 S2蛋白、HIV-1 gp41蛋白前述高度(构造)相似性的论文,丝毫未提及其它冠状病毒S2蛋白与HIV-1 gp41蛋白的相似性。
上面指出的是SARS-CoV-1、HIV-1融合蛋白间的高度构造相似性。那么,SARS-CoV-2是否继承、保持了SARS-CoV-1 S2蛋白(S2亚基)与HIV-1 gp41蛋白之间的高度构造相似性呢?
1)SARS-CoV-2 、SARS-CoV-1 S2蛋白有很高的序列相似性。二者Spike蛋白S1亚基aa序列一致性仅为64.85%,但是,S2亚基一致性高达90.10%;
2)SARS-CoV-2、SARS-CoV-1 S2亚基共有高度序列相似的两个七肽重复区域(N-HR1、C-HR,也叫HR1、HR2),以及C-HR和跨膜区域之间的富含芳香族残基区域。二者N-HR(HR1)区域的aa序列一致性约为92.05%,二者这一区域的88个氨基酸残基中的前13个部分相同,后75个完全相同;二者C-HR(HR2)区域+富含芳香族残基区域的aa序列一致性约为98.08%,二者这两个区域的共52个残基仅有一个不同。
3)SARS-CoV-2全部保持了SARS-CoV-1 N-HR(HR1)、C-HR(HR2)间环结构中的4个半胱氨酸(C)残基、9个脯氨酸残基(P),二者两个HR间环结构区域的aa序列一致性约为92.76%;
由此推断,SARS-CoV-2 S2亚基应该继承、保持了SARS-CoV-1 S2亚基与HIV-1 gp41亚基的高度构造相似性。
注:上述比对结果所用SARS-CoV-2 Spike蛋白genbank accession number:YP_009724390.1/1796318598,对应Wuhan-hu-1毒株;SARS-CoV Spike蛋白genbank accession number:AAP13441.1,对应SARS Urbani毒株。
HIV-1、SARS-CoV-1、SARS-CoV-2的膜融合机制也高度相似:都通过两个七肽重复区域N-HR、C-HR的相互作用、重新折叠形成六螺旋束(six-helical bundle,6-HB)结构,六螺旋束将病毒包膜与细胞膜/内体膜/溶酶体膜拉近,从而导致膜融合。不过,这一膜融合机制并非三种病毒专有,它应该为I类包膜病毒(type I enveloped viruse)所共有。I类包膜病毒包含许多高致病性逆转录病毒、冠状病毒、流感病毒、副粘病毒和丝状病毒,它们的融合蛋白为I类融合蛋白。
通过在S1/S2处(S1、S2亚基交界处)插入furin酶切位点(furin cleavage site,FCS),SARS-CoV-2优化了SARS-CoV-1的膜融合、感染、复制过程。由于存在S1/S2处的FCS,完成受体结合后,SARS-CoV-2的Spike可在细胞外被furin蛋白酶水解切割,暴露S2亚基,激活其膜融合活性,介导病毒包膜直接与细胞膜融合,第一时间将病毒包膜内的病毒RNA释放到细胞内(细胞质中),启动病毒复制。
SARS-CoV-2 Spike蛋白、HIV-1 gp160蛋白都具有FCS,二者的FCS都恰好位于Spike/gp160两个亚基的交界处,SARS-CoV-2、HIV-1都可通过直接膜融合达成高效感染。
尽管膜融合蛋白构造、膜融合机制与HIV-1高度相似,但SARS-CoV-1的Spike没有FCS,不能在细胞外被水解切割,它的病毒包膜不能与细胞膜直接发生膜融合。完成受体结合后,SARS-CoV-1将被整体内吞,其Spike将在细胞内被组织蛋白酶B或L(CatB/L)水解切割,而后,病毒包膜再与内体膜或溶酶体膜发生膜融合以释放病毒RNA,启动病毒复制。SARS-CoV-1内吞+细胞内膜融合的感染效率远低于SARS-CoV-2/HIV-1细胞外直接膜融合的感染效率,SARS-CoV-2的感染效率约是SARS-CoV-1的100~1000倍。
3、SARS-CoV-2继承了SARS-CoV的2个肉毒杆菌神经毒素样基序,1个人类ICAM-1样基序。
xxx样基序(xxx-like motif),指与xxx aa序列(amino acid sequence,氨基酸序列)相似,对应的多肽链空间结构也相似并具有共同生物功能/能力的基序,基序,可理解为具有特定功能的一个aa序列片断或一组邻近的aa序列片断。肉毒杆菌神经毒素样基序,即与肉毒杆菌神经毒素(Botulinum neurotoxin)aa序列相似,多肽链空间结构相似,并具有肉毒杆菌神经毒素生物功能/能力的序列片断。
SARS-CoV-2 Spike蛋白含有2个肉毒杆菌神经毒素样基序(简记为neurotoxin-like motif),S1亚基、S2亚基内各有一个。其中,位于S1亚基内,对应Spike蛋白T299 ~Y351序列片断,与G型肉毒杆菌神经毒素前体(Botulinum neurotoxin type G precursors)相似的基序能够强力结合TCR(T cell receptor,T细胞受体),并触发强烈的T细胞免疫反应。非常巧合的是,SARS-CoV-2 Spike中的这2个neurotoxin-like motifs在SARS-CoV-1 Spike中的对应序列同样也是neurotoxin-like motif,2对基序的一致度分别为75%、84%。这可以解读为,SARS-CoV-2 继承了SARS-CoV-1的2个neurotoxin-like motifs,并对其作了某些调整、优化。SARS-CoV-1 Spike共有3个neurotoxin-like motifs,它的另一个基序在SARS-CoV-2 Spike中的对应序列不是neurotoxin-like motif。
SARS-CoV-2 Spike蛋白S1亚基还含有一个ICAM-1样基序(ICAM-1-like Motif),该基序同样具有TCR结合能力,它通过结合TCRVα(Variable α-domain of TCR,TCR的可变α 域)增进、稳固了neurotoxin-like motif(T299 ~Y351)与TCR的结合。neurotoxin-like motif(T299 ~Y351)可能同时结合TCRVα和TCRVβ(Variable β-domain of TCR,TCR的可变β域)。ICAM-1(Intercellular Adhesion Molecule-1,细胞间粘附分子-1 ),也叫做CD54 (Cluster of Differentiation 54,分化簇54),是一种细胞表面跨膜糖蛋白,通常在内皮细胞和免疫系统细胞的细胞膜上表达,对于免疫和炎症反应的介导至关重要。ICAM-1也在中枢神经系统(central nervous system,CNS)中表达,如在人类前脑的白质和灰质中的小胶质细胞、星形细胞以及内皮细胞中表达。ICAM-1在血脑屏障(blood-brain barrier)中具有关键作用,在精神疾病的发病机制中也有重要作用。
SARS-CoV-2 Spike中的ICAM-1-like Motif在SARS-CoV-1 Spike中的对应序列也是ICAM-1-like Motif,二者一致度为74%。SARS-CoV-1 Spike含有2个ICAM-1-like Motif。

SARS-CoV-2/SARS-CoV-1 Spike中的neurotoxin-like motifs和ICAM-1-like motifs
图a:SARS-CoV-2/SARS-CoV-1 Spike中的neurotoxin-like motifs(绿色行)、ICAM-1-like motifs(橙色行)的有关信息;
图b:SARS-CoV-2 Spike三聚体(trimer)带状模型一个单体(monomer,黄色)上的两个neurotoxin-like motifs(绿色圆圈指示)、一个 ICAM-1-like motif(橙色圆圈指示)及S1/S2处的PRRA插入region(紫色圆圈指示);
图c:SARS-CoV-2 Spike-TCR复合体、SARS-CoV-1 Spike-TCR复合体填充模型中与TCR结合的neurotoxin-like motifs。
SARS-CoV-2 Spike蛋白新增了1个SEB样基序(SEB-like motif),该基序同样具有强大的TCR结合能力。SEB(Staphylococcal enterotoxin B,葡萄球菌肠毒素B,肠毒素B)是金黄色葡萄球菌的代谢、分泌产物,是一种强大的超级抗原(Superantigen,SAg)毒素,是公认的毒素类生物、化学战剂,生物恐怖制剂。SARS-CoV-1 Spike中没有SEB-like motif。
SARS-CoV-2 Spike蛋白中具有SAg能力的肠毒素B样基序(SEB-like motif)和肉毒杆菌神经毒素样基序(neurotoxin-like motifs)可能是COVID-19重症患者T细胞大规模激活、增殖,引发细胞因子风暴,造成多器官损伤的关键因素(之一)。
SARS-CoV-2 Spike的neurotoxin-like motifs、ICAM-1-like motif、SEB-like motif相辅相成,共同服务于以下致病链条:TCR结合—T细胞过度激活--诱发细胞因子风暴、免疫风暴、过度炎症反应--造成多器官损伤;上述3类基序可能也参与SARS-CoV-2 对大脑、神经系统的感染、损伤;SARS-CoV-2 还能通过结合CD4(、TCR)感染、破坏CD4+T淋巴细胞。CD4、CD8是TCR的两种协同受体,CD4+T淋巴细胞是表面附着CD4的淋巴细胞。
SARS-CoV-2 Spike还含有一个蛇神经毒素样区域/基序(snake-toxin like region),该区域/基序与眼镜蛇属和银环蛇属神经毒素同源(序列显著相似)。
4、SARS-CoV-2继承、优化了SARS-CoV nsp3蛋白的SARS-unique domain。
Nsp3(non-structural protein 3,非结构蛋白3)是冠状病毒基因组编码的最大蛋白质,包含至少8个功能模块/结构域,对病毒复制、病毒毒力至关重要。SARS-CoV-2、SARS-CoV的nsp3都含有SARS独特结构域(SARS-unique domain,SUD),拥有SUD是这两种病毒高致病力的关键决定因素之一。

SARS-CoV nsp3组件构成及SUD 3D结构的带状模型
SARS-CoV nsp3的SUD包含三个子域(subdomains):SUD-N(N-terminal subdomain)、SUD-M(middle subdomain)、SUD-C(C-terminal subdomain)。这三个SUD子域在SARS-CoV-2 nsp3中同样存在。

SARS-CoV-2 nsp3组件构成

SARS-CoV-2、SARS-CoV SUD的aa序列比对及结构模型(未显示SUN-C)对照
SUD三个子域中的SUD-N、SUD-M合称SUD-core,这两个SUD子域以及SUD之前的X 结构域(X domain,Mac1)都是能够抑制宿主蛋白抗病毒活性的宏结构域(macrodomain),这三个nsp3中相邻的宏结构域也依次被称为Mac1、Mac2、Mac3。同时拥有三个宏结构域,可能是SARS-CoV-2、SARS-CoV在抑制、对抗宿主抗病毒反应方面比其他(冠状)病毒更有效的重要原因。绝大多数冠状病毒只具有1~2个宏结构域。
SARS-CoV-2、SARS-CoV SUD的aa序列一致度约为75%。二者nsp3中的SUD相似但不雷同,各有短长;相比SARS-CoV SUD,SARS-CoV-2 SUD在解决方案上具有高度灵活性。
完整的SUD是β谱系B支系(乙型冠状病毒属B亚属)某些冠状病毒的特有结构。2009年的一篇论文指出:SUD在除SARS-CoV外的所有冠状病毒中都不存在,具有SUD可能是SARS-CoV具有极高致病性的原因;2015年的一篇论文发现,MHV(Mouse hepatitis virus,小鼠肝炎病毒)nsp3中的DPUP(Domain Preceding Ubl2 and PL2pro)结构域与SARS-CoV SUD的SUD-C子域具有密切的结构相似性。MHV是属于β谱系A支系的一类冠状病毒;同年另一篇论文发现,MERS-CoV及MERS-related CoV(HKU4、 HKU5)nsp3含有与SUD-M相似的残基簇(cluster of residues),还有论文指出,MERS-CoV中也存在与SUD-C相似的结构域。MERS-CoV及MERS-related CoV属于β谱系C支系;然而,β谱系B支系之外、β谱系之外的冠状病毒无一拥有完整的SUD。含有SUD-N,并完整拥有SUD-N、SUD-M、SUD-C三个SUD子域的冠状病毒只存在于β谱系B支系中,可能只有SARS-CoV和SARS-CoV-2。有些论文指出,β谱系B支系的某些SARS-like蝙蝠冠状病毒,如WIV1、WIV16、Rs3367、RsSHC014也具有SUD,但这些论文并没有明确指出、显示这些蝙蝠冠状病毒拥有完整的SUD。上述四种病毒都是具有ACE2结合能力,甚至具有人体细胞进入能力的特殊蝙蝠冠状病毒,但是,它们都没有人类致病能力,都不会使人体产生临床疾病症状。
SARS-CoV、SARS-CoV-2 SUD可结合多种宿主蛋白,以及具有G4结构的宿主细胞DNA、RNA或病毒自身RNA(SARS-CoV-2的SUD可能不能结合自身RNA中的G4)。
SARS-CoV、SARS-CoV-2 SUD(full-length SUD及子域SUD-N和SUD-M)都能结合DNA/RNA中的鸟嘌呤四链体(guanine quadruplexes,G-四链体,G4),它们通过结合含有G4的宿主细胞RNA/mRNA来调节、促进病毒复制、转录,并干扰、对抗宿主细胞的抗病毒免疫反应。G4是一种超螺旋(superhelix)结构,存在于DNA 或RNA 链中富含G 碱基(鸟嘌呤)的区域内,是4条鸟嘌呤核酸链通过Hoogsteen 型氢键(Hoogsteen-type hydrogen bonds)配对互联形成。G4广泛参与人和动物的DNA 复制、转录、翻译和端粒结构维持等一系列基本生物学功能。有论文指出,SARS-CoV-2 SUD-core中松散连接的SUD-N和SUD-M在与G4 结合时具有更大的灵活性,而SARS-CoV SUD-core中连接SUD-N、SUD-M的二硫键桥(disulfide bridge)限制了SUD-N、SUD-M在G4 结合过程中的构象变化。

SARS-CoV-2、SARS-CoV SUD-G4作用、结合机制对比图
SARS-CoV-2、SARS-CoV SUD都能作用、结合人类Paip1,并通过结合Paip1来利用/劫持/霸占细胞的转录、翻译机制,刺激、促进、增强病毒翻译和病毒蛋白合成,但不促进宿主细胞mRNA翻译及宿主蛋白合成。Paip1(PABP-interacting protein 1,PABP相互作用蛋白1)是一种宿主翻译因子-蛋白质翻译刺激剂,人类Paip1 有三种亚型(isoforms);PABP(Poly(A)-binding protein,多聚腺苷酸结合蛋白)是一类可干预生物体/病毒转录、翻译的生物体蛋白质,PABP 可与生物体自身mRNA或病毒mRNA3'端多腺苷酸尾结合,保护所结合的mRNA不被降解。人类和脊椎动物有五种PABP。PABP 和 Paip1 都是人体细胞翻译起始复合体(translation initiation complex)的组成部分,SARS-CoV-2、SARS-CoV SUD与Paip1的结合会增强Paip1与PABP 的结合亲和力,SUD可能与Paip1、PABP形成三元复合体。

SARS-CoV-1 SUD-N与Paip1M作用、结合示意图
图A左:SUD-N(orange)、Paip1M(light blue)复合物晶体结构的带状模型;
图A右:SUD-N(orange)、Paip1M(light blue)复合物晶体结构的空间填充模型。
Paip1M是Paip1的中间域(the middle domain of Paip1)。SARS-CoV-2、SARS-CoV SUD都以子域SUD-N与Paip1M作用结合。

SARS-CoV-2 SUD(框中彩色条带)、Paip1(灰色条带)复合物的带状模型
SARS-CoV-1 SUD还协同nsp3的另一组件—PLpro/PL2pro(papain-like protease,木瓜蛋白酶样蛋白酶。SARS-CoV-1、SARS-CoV-2中的PLpro是PL2pro,即木瓜蛋白酶样蛋白酶2)结构域与E3 泛素连接酶(E3 ubiquitin ligase)--RCHY1相互作用、结合,劫持、利用RCHY1介导泛素化(ubiquitination)和人类p53蛋白降解,抑制宿主细胞/p53蛋白的抗病毒活性,从而促进病毒复制。不包含SUD或不包含完整SUD的的病毒,如MERS-CoV、HCoV-NL63,也能通过PLpro/PL2pro 域结合RCHY1,促使p53蛋白降解;相比之下,SARS-CoV-1的SUD-PLpro 融合物(SUD–PLpro fusion)与RCHY1的结合更强烈,p53蛋白降解效果更显著(导致更多的p53蛋白被降解)。
尽管未经公开发表的论文实验验证,但有论文暗示,这是SARS-CoV-1与SARS-CoV-2的共有能力,即SARS-CoV-2的SUD、PLpro/PL2pro 也能强烈结合RCHY1,促使p53蛋白显著降解。另外一篇探讨SARS-CoV-2致癌性的短论文则明确认为,SARS-CoV-2兼具SARS-CoV-1 的上述能力。
RCHY1(RING finger and CHY zinc finger domain-containing protein 1,包含环指和CHY-type锌指结构域的蛋白1)是一种具有泛素连接酶活性的人类蛋白,该蛋白可连接/结合p53蛋白并导致后者被降解(水解)。p53蛋白是一种重要的多功能人体蛋白,其功能之一是作为抗病毒因子抑制病毒复制。泛素连接酶/E3泛素连接酶,是能够将泛素分子连接到目标蛋白质的某个赖氨酸上的酶。泛素(ubiquitin)是一种存在于大多数真核细胞中的小蛋白,它的主要功能是连接、标记需要分解的蛋白质,相应蛋白酶可识别被泛素附着、标记的蛋白质,并将被标记的蛋白质水解。
SARS-CoV-2、SARS-CoV SUD通过结合RCHY1促使p53蛋白降解这一能力不仅促进病毒复制,而且具有高度致癌性。p53蛋白也被称为p53肿瘤蛋白/p53肿瘤抑制蛋白,是一种重要的肿瘤抑制因子,p53蛋白突变或功能丧失将导致高度的细胞癌变风险,p53蛋白的长期减少同样是致癌的重大危险因素。
SARS-CoV-2与p53蛋白存在多重关联。SARS-CoV-2的Spike蛋白等蛋白含有与p53蛋白相似的p53结构域(p53 domain),可能模仿、介入、干预、干扰p53蛋白的生理、病理功能;SARS-CoV-2 Spike蛋白S2亚基的HR-2(第二个七肽重复区域,即前文所说的C-HR)能够直接地强烈作用、结合p53蛋白及另外两种肿瘤抑制蛋白--BRCA-1/2(BReast CAncer gene-1/2,乳腺癌基因-1/2)蛋白,这意味着SARS-CoV-2 Spike具有强大的p53蛋白劫持能力。

SARS-CoV-2 S2 subunit HR-2与p53、BRCA-1、BRCA-2 strongly interact示意图
SARS-CoV-2的另一种non-structural protein--nsp2与主要位于细胞线粒体中的两种抑制素(prohibitin)PHB1、PHB2相互作用,使活性氧(reactive oxygen species,ROS)泄漏到细胞核产生氧化损伤,导致合成p53蛋白的依赖基因被破坏。活性氧(ROS)失调、紊乱也是重要的致癌因素。
SARS-CoV-2与p53蛋白的多重关联只是SARS-CoV-2高度致癌性的一组因素,SARS-CoV-2具有非常多样化的致癌途径。
SARS-CoV-2对人类p53蛋白的间接降解、模仿/干扰、直接结合/劫持,以及多途径、系统性的人类致癌机制,是在动物宿主中发生的,无目的、随机性自然变异/自然演化的结果与巧合吗?
5、SARS-CoV-2继承、拓展了SARS-CoV E蛋白、ORF3a蛋白的PDZ结合基序。
SARS-CoV-2、SARS-CoV E蛋白(Envelope protein)、ORF3a蛋白的C-末端(C-terminal,COOH端,羧基端)都含有PDZ结合基序(PDZ-binding motif,PBM)。

SARS-CoV E蛋白C末端的PBM

SARS-CoV-2 E蛋白C末端的PDZ-binding motif
PBM是符合特定组合模式的短基序,有三种类型。SARS-CoV-2、SARS-CoV E蛋白C末端的PBM同为DLLV(天冬氨酸-亮氨酸-亮氨酸-缬氨酸),是一个type-II PBM。
SARS-CoV-2、SARS-CoV E蛋白高度相似,二者E蛋白aa序列相似度为94.7%;二者ORF3a蛋白虽然差异较大(aa序列相似度仅为73.1%),但它们的C末端也都含有PBM。
病毒蛋白C末端的PBM是病毒毒力/致病能力的另一重要决定因素。
PDZ-binding motif(PBM)与PALS1(Protein Associated With Lin Seven 1)相互作用、结合并改变后者的亚细胞定位,拉扯PALS1,使其脱离原有的细胞内位置,导致感染者上皮形态改变,使原本致密的组织结构撕裂、破损、渗漏,造成上皮组织(如肺上皮组织)的广泛损伤及其它组织损伤。上皮损伤、渗漏、屏障功能丧失后SARS-CoV 病毒将渗入底层组织,并通过体循环扩散到其它组织、器官,造成更广泛的感染、损伤、破坏。PALS1是一种衔接细胞内组件的鸟苷酸激酶蛋白,PALS1蛋白/基因存在于心脏、肝脏、肾脏、肺、肠、血管、神经系统、眼睛、肌肉、皮肤等众多器官、组织中。PALS1内的PDZ domain是SARS-CoV-2/SARS-CoV PBM(PDZ-binding motif)的结合位点。
PBM还结合另一种含有PDZ domain的人类细胞蛋白Syntenin(内居蛋白),并以此激活p38 MAPK(p38 mitogen-activated protein kinase,p38丝裂原活化蛋白激酶),导致炎症细胞因子过度表达,诱发“细胞因子风暴”。p38 MAPK是一类细胞内信号传导因子,参与炎症调节、氧化应激、细胞凋亡等生理、病理过程。PBM结合PALS1造成组织损伤后,免疫细胞试图修复损伤时释放细胞因子,也会诱发或加剧大规模炎症及“细胞因子风暴”。
PDZ即PSD-95/Dlg/ZO-1,是三种蛋白首字母的缩写:Post synaptic density protein(突触后密度蛋白,PSD-95)、Drosophila disc large tumor suppressor(果蝇盘大肿瘤抑制蛋白,Dlg1)和Zonula occludens-1 protein(小带闭塞-1蛋白,zo-1)。PDZ domain当初被发现是以上三种蛋白的共有结构,因而得名,后来发现,PDZ domain存在于大量人类和动物蛋白之中,可介导众多人类蛋白(/动物蛋白)相互作用。病毒蛋白中的PBM(PDZ-binding motif)可结合具有(相应?)PDZ domain的人体/动物蛋白。
截止2020年,共有7种已知的人类致病冠状病毒。除SARS-CoV-2、SARS-CoV外,其它5种病毒(MERS-CoV、HCoV-OC43、HCoV-HKU1、HCoV-NL63 和HCoV-229E)E蛋白(C末端)也含有PBM,但已论文报告的,ORF3a蛋白(C末端)含有PBM的冠状病毒可能只有SARS-CoV-2和SARS-CoV,ORF3a蛋白对SARS-CoV-2的致病能力同样非常重要。在SARS-CoV基础上,SARS-CoV-2还更进一步,除E蛋白、ORF3a蛋白外,它的N蛋白(Nucleocapsid protein,核衣壳蛋白)的C-末端也含有PBM。
SARS-CoV-2 PBM的结合对象远不只PALS1、Syntenin两种。研究表明,SARS-CoV-2 E、ORF3a和N蛋白中的PBM至少可结合16种人类PDZ结合物(human PDZ binders),其中6种结合物也可被SARS-CoV(E、ORF3a蛋白)的PBM结合。16种SARS-CoV-2人类PDZ 结合物中的大多数是参与细胞衔接/连接或维持细胞极性的人体蛋白,SARS-CoV-2可能通过结合、劫持这些细胞元件来服务于自身的感染、复制、致病和免疫逃避。
6、SARS-CoV-2的铁调素模仿能力、铁转运蛋白结合能力可基于SARS-CoV简单改造获得。
SARS-CoV-2 Spike蛋白尾部区域能够模仿人类铁调素(human Hepcidin),具备铁调素的铁转运蛋白结合等能力。铁转运蛋白(ferroportin,FPN,又称膜铁转运蛋白)是已知的唯一一种细胞铁输出蛋白;铁调素与铁转运蛋白的结合具有特异性,铁调素可能是已知的唯一一种/类能够结合铁转运蛋白的天然生物蛋白或多肽。
具有铁调素模仿能力、铁转运蛋白结合能力的,是SARS-CoV-2 Spike蛋白含26个氨基酸的尾部区域,SARS-CoV-2的这一区域与SARS-CoV的对应区域高度相似。

8种冠状病毒Spike尾部区域,及4种铁调素的aa序列比对图
如上图所示,SARS-CoV-1、SARS-CoV-2对应区域内的26个氨基酸仅有两个不同:第3位点的L(亮氨酸)~M(甲硫氨酸),第17位点的A(丙氨酸)~C(半胱氨酸)。基于SARS-CoV-1的对应区域,将区域内第3位点的L(亮氨酸)替换为M(甲硫氨酸),将第17位点的A(丙氨酸)替换为C(半胱氨酸),即可获得SARS-CoV-2的铁调素模仿、铁转运蛋白结合区域。
笔者不确定SARS-CoV(尾部区域)是否也具有人类铁调素模仿、铁转运蛋白结合能力,当前没有论文研究、检验、提及SARS-CoV的相应能力。本文假定SARS-CoV不具备这组能力。SARS-CoV-2可能是已论文报告的,唯一一种具有人类铁调素模仿能力的病毒,唯一一种可结合人类铁转运蛋白的病毒。SARS-CoV-2的Spike还具有人类铁调素结合能力,它可能也是已论文报告的,唯一一种可结合人类铁调素的病毒。
相关文章:
万能”受体适配器--新冠S蛋白 4
7、SARS-CoV-2的DPP4结合能力可基于SARS-CoV简单改造获得。
SARS-CoV-2兼具MERS-CoV的DPP4结合能力,并可以DPP4为受体感染某些人体细胞,如脑和脊髓中的人皮质星形胶质细胞。SARS-CoV-2 Spike蛋白RBD中决定DPP4(dipeptidyl peptidase IV,二肽基肽酶4,也叫CD26)结合能力的4个关键氨基酸残基是:D405(天冬氨酸)、E484(谷氨酸)、Y489(酪氨酸)、Q498(谷氨酰胺)。

SARS-CoV-2的4个DPP4结合关键氨基酸及SARS-CoV中的对应氨基酸
由上图可见,4个关键氨基酸中的D405、Y489在SARS-CoV中对应相同的氨基酸;而E484
、Q498在SARS-CoV中对应不同的氨基酸:E484在SARS-CoV-1中的对应位点处空缺了一个氨基酸,Q498
在SARS-CoV-1中的对应位点是一个酪氨酸(Y)。
在SARS-CoV-1的E484对应位点处插入一个谷氨酸(E),并将Q498对应位点处的酪氨酸(Y)替换为谷氨酰胺(Q),就基于SARS-CoV-1
Spike获得了SARS-CoV-2 Spike的DPP4结合能力。SARS-CoV Spike不能有效结合DPP4。
相关文章:
万能”受体适配器--新冠S蛋白 1
8、SARS-CoV-2改进了对HS/HSPG结合、利用。
HS/HSPG是SARS-CoV-2的众多感染受体之一,它对SARS-CoV-2感染的介导依赖ACE2,是ACE2的辅助受体或共受体;SARS-CoV-1也有一定程度的HS/HSPG结合能力,但它应该不是SARS-CoV-1的感染受体,有研究表明,HS/HSPG干扰、阻碍而非促进SARS-CoV-1感染。
HS:heparan sulfate,硫酸乙酰肝素,是一种人类和哺乳动物组织中普遍存在的,带高负电荷的、硫酸化的链状糖胺聚糖;
HSPG:heparan sulfate proteoglycans,硫酸乙酰肝素蛋白聚糖,是1~3个HS多糖链与某个蛋白通过共价键连接形成的复合体。
SARS-CoV-2的HS/HSPG结合能力强于SARS-CoV。
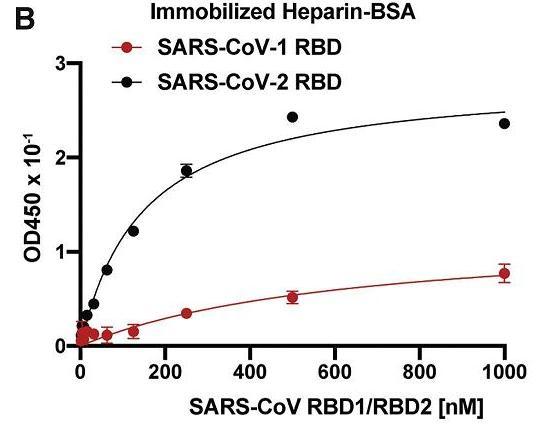
SARS-CoV-2、SARS-CoV-1 RBD的HS/Heparin结合能力对比
相对SARS-CoV-1,SARS-CoV-2 HS/HSPG结合能力的显著提升,大致可通过两个氨基酸替换(Glu354Asn、Thr444Lys)/(E354N、T444K)获得。
SARS-CoV-2 Spike中对结合HS/HSPG贡献较大的氨基酸残基有8个,这8个氨基酸及它们在SARS-CoV-1中的对应位点氨基酸在下图中用红色方框标出。

SARS-1、SARS-2中决定HS/HSPG结合能力的重要氨基酸残基
8个HS/HSPG结合重要氨基酸中的4个在SARS-CoV-1 中对应相同的氨基酸,另外4个对应不同的氨基酸,4对差异氨基酸(残基)是:K346R、K357R、E354N、T444K。氨基酸单字母缩写中间的数字是后一个氨基酸残基在SARS-CoV-2 Spike aa序列中的序号。
4对差异氨基酸中的前两组K346R、K357R都是赖氨酸K到精氨酸R的保守替换。赖氨酸、精氨酸都是碱性残基,二者物理、化学性质接近,它们之间的替换是保守替换,对HS/HSPG结合能力的影响不大;对HS/HSPG结合能力提升起关键作用的是另外两组差异氨基酸:E354N、T444K,用三字母缩写表示即:Glu354Asn、Thr444Lys。
E、Glu是谷氨酸的单字母缩写、三字母缩写;
N、Asn是天冬酰胺的单字母缩写、三字母缩写;
T、Thr是苏氨酸的单字母缩写、三字母缩写;
K、Lys是赖氨酸的单字母缩写、三字母缩写。
SARS-CoV-2 Spike蛋白的第354位点对应SARS-CoV-1 Spike蛋白的第341位点,SARS-CoV-2 Spike第444位点对应SARS-CoV-1 Spike的第431位点。至此可知,以天冬酰胺(Asn,N)替换SARS-CoV-1 Spike第341位点的谷氨酸(Glu,E),同时,以赖氨酸(Lys,K)替换SARS-CoV Spike第431位点的苏氨酸(Thr,T),简单改造后的SARS-CoV-1 Spike就获得了与SARS-CoV-2 Spike大体相当的HS/HSPG结合能力。
相关文章:
万能”受体适配器--新冠S蛋白 2
9、其它受体关联
除了都以ACE2为(首要)感染受体外,SARS-CoV-2、SARS-CoV-1还至少共有另外4种感染受体:CD147/Basigin(基础免疫球蛋白)、vimentin(波形蛋白)、CD209L/L-SIGN、CD209/DC-SIGN。
CD147/Basigin
可以不依赖ACE2独立介导SARS-CoV-2感染,如介导SARS-CoV-2对T细胞(T淋巴细胞)的感染,也可与ACE2共同介导SARS-CoV-2感染,如介导对肾脏足细胞(肾小球上皮细胞)的感染。vimentin、CD209L/L-SIGN、CD209/DC-SIGN对SARS-CoV-2/SARS-CoV-1感染的介导应该都依赖于ACE2,是ACE2的辅助受体/共受体。
问题
巧合太多了就不再是巧合了。
2005年之后,SARS-CoV就在实验室外销声匿迹了。如果SARS-CoV-2是自然产生的,那么,它从何种祖先身上获得(继承或重组)了如此之多的SARS-CoV基因结构?如果SARS-CoV-2来自动物,那么,未经任何适应性突变的SARS-CoV-2,何以刚刚接触人类就以人类为最佳感染对象?就拥有如此深思熟虑、周祥完备、精雕细琢、游刃有余的人类感染、致病机制?这些、这样的人类感染、致病机制可能在蝙蝠、穿山甲或其它动物宿主体内演化产生吗?它们都是无目的、随机性自然变异、自然进化、自然重组、自然演化的结果、巧合吗?
相关文章:
“万能”受体适配器--新冠S蛋白 5 上
谁设计、制造了新冠病毒(四)匪夷所思
(未完待续)